Resesif, dominant, öldürücü genler, mutasyonlar. III.2.8
Şimdiye kadar insan genetiğini kalıtım türlerine göre ele aldık; şimdi genlerin çok önemli bir kategorisine, yani öldürücü ve öldürücü olmayan genlere döneceğiz. Ölümcül ve yarı öldürücü genler, gelişim üzerinde belirgin olumsuz etkileri olan bir mutasyon sınıfıdır. Daha sonra göreceğimiz gibi, çoğu durumda mutasyonlar kişinin kalıtımını kötüleştirir ve yok eder. Bu


Mutasyonların şiddetlenmesi özellikle ölümcül genlerin etkisi altında belirgindir.
Ölümcül ve öldürücü olmayan aleller baskın veya resesif olabilir. Ölümcüllüğün baskın olması durumunda çocuk, doğum öncesi dönemde veya erken bebeklik döneminde ölür. Bu tür genlerin kalıtsal olarak aktarılamayacağı oldukça açıktır. Onların varlığı mutasyonların ortaya çıktığını gösterir. Bu tür baskın öldürücülerin toplam sıklığı o kadar da küçük değil. Radyasyonun etkisi altında sayıları özellikle fazladır. Farelerin ve Drosophila'nın genetiğinde, mutasyon sıklığının doza bağımlılığını analiz ederken ve mutajenik etkilerin diğer özelliklerini analiz ederken baskın öldürücüler yaygın olarak kullanılmaktadır. Yarı öldürücü genler olarak da adlandırılan alt öldürücü genlerin varlığında, çocuk erken çocukluk döneminde veya bir süre sonra, ancak kural olarak ergenliğe ulaşmadan önce ölür. Baskın yarı ölümcül mutasyonun bir örneği, negatif semptomların bütün bir sendromunun etkisiyle karakterize edilen bir hastalık olan epiloiadır. Epilojili çocuklarda ciltte patolojik büyümeler vardır; zeka geriliği, epilepsi, kalp, böbrek ve diğer organ tümörleri ile karakterize edilirler. Nadir durumlarda epiloili hastalar hayatta kalır ve sağlıklı bir eşle evlenir. Böyle bir evlilikten doğan çocukların yarısı konjenital epiloya hastasıdır (Şekil 131).
Ölümcül olmayan dominant mutasyonun bir başka çarpıcı örneği de retinoblastoma hastalığıdır. Bu


Baskın gen gözde kanserli bir tümöre neden olur ve bu da neredeyse her zaman çocuğun erken yaşta ölümüne yol açar.
Bununla birlikte, konjenital deformitelerin büyük kısmı resesif öldürücü ve yarı öldürücü türlerden kaynaklanmaktadır. Bu durumda, görünüşte sağlıklı ebeveynlerin genotipinde resesif zararlı genler gizlidir, ancak bu tür iki heterozigot evlendiğinde çocuklarının yaklaşık %25'inin hasta olduğu ortaya çıkar.
Şekil 132, resesif öldürücü ve öldürücü olmayan genlerin miras alınması sırasında insan popülasyonlarında gerçekleştirilen ana melezleme türlerini göstermektedir. Kare 1 genetik olarak normal insanlar arasındaki evlilik türlerini sunar. Hem ebeveynlerde hem de onların soyundan gelenlerin tümünde, tüm aleller normal genler (alel +) ile temsil edilir. Kare 2 genetik olarak sağlıklı bir kişi ile bir heterozigot arasındaki evliliği tasvir eder (sembol a). Bu durumda, negatif alel yavruların yarısına aktarılır, ancak gizli kalır. Kare 3 genetik olarak sağlıklı bir kişi hasta bir kişiyle evlenir (sembol aa), Görünüşte tüm çocuklar sağlıklıdır ancak heterozigot durumdayken ölümcül olmayan bir mutasyon taşırlar. A. Kare 4 bir heterozigot bir hastayla evlenir; çocuklarının yarısı görünüşte sağlıklı ama ölümcül olmayan bir mutasyon taşıyor, diğer yarısı ise hasta. Son olarak bir kare 5 iki hasta kişinin evlenmesi durumunda (semboller aa) tüm yavrular hasta olacak. Resesif öldürücü ve öldürücü olmayan genlerin etkisinin neden olduğu konjenital hastalıklar arasında beyin dokusunun dejenere olduğu, körlüğün meydana geldiği ve çocuğun öldüğü çocukluktaki amatör aptallık; varlığı ile konjenital cilt patolojisi
derin kanama çatlakları - iktiyoz (Şekil 133); konjenital çocuk felci ve bir dizi başka konjenital hastalık.
Erken dönem spontan düşüklerin çoğu, ölümcül genotiplerin sızmasından kaynaklanmaktadır.
Baskın mutasyonların, heterozigotlarda az çok nötr bir özelliği belirlediği, ancak homozigot durumda öldürücü bir etkiye sahip olduğu bilinmektedir. Örneğin, heterozigot durumdaki kseroderma pigmentasyon aleli ciddi çillenmeye neden olur. Ancak homozigotlarda kseroderma pigmentosum gelişir. Bu, ışığın etkisinin ciltte hasara neden olduğu ve daha sonra kötü huylu büyümeye yol açtığı ölümcül olmayan bir hastalıktır (Şekil 134).
Heterozigot durumdaki bir kişideki alellerden biri kanda aşırı kolesterole neden olur. Bununla birlikte, bu alel için homozigot olan çocuklarda ciddi, ölümcül olmayan bir anormallik gelişir.
Tüm bu durumlarda, daha önce Drosophila, fareler ve diğer organizmalarla yapılan deneylerde ortaya konmuş, resesif öldürücü etkiye sahip dominantların kalıtımına ilişkin klasik bir tabloyla karşı karşıyayız. Örneğin farenin iyi bilinen bir aleli vardır. Bir E, bu da heterozigotlarda derinin sararmasına neden olur. Ancak saf bir sarı fare ırkı elde etmeye yönelik tüm girişimler başarısızlıkla sonuçlandı. İki sarı fareyi çaprazlarken, ortalama olarak iki sarı fare her zaman farklı renkte bir tane, örneğin siyah üretir. Bu olgunun doğası, baskın sarı renk geninin ortaya çıkmasıyla açıklandı.
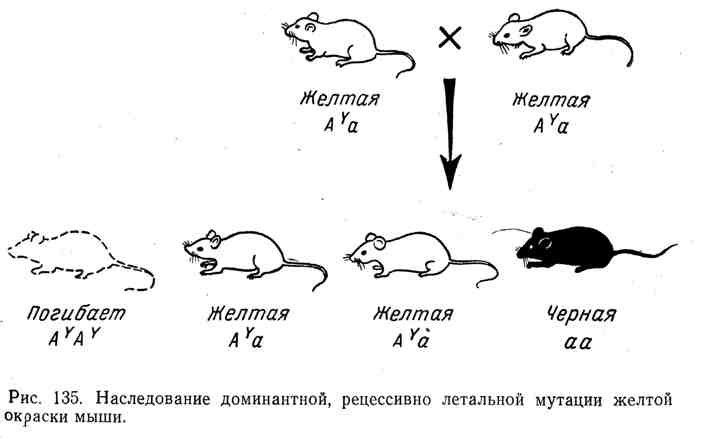
resesif öldürücü. Tüm homozigotlar( AY AY) ölü. Sarı fare alel ile birlikte her zaman heterozigotturBir E başka bir alel içerir; A(siyah renk). Heterozigotları geçerken kalıtımın seyri A Ya ve 2:1 formülüne göre bölme Şekil 135'te gösterilmektedir.
- Kaynak-
Dubinin, N.P. Genetiğin ufukları / N.P. Dubinin. – M.: Eğitim, 1970.- 560 s.
Gönderi Görüntüleme Sayısı: 121
Farklı genotiplerdeki zigotların eşit olmayan canlılığı, ebeveyn bireylerin gametlerinde ortaya çıkan baskın veya resesif öldürücü mutasyonlarla ilişkilendirilebilir. Embriyogenezin farklı aşamalarında veya embriyonik sonrası dönemde gerçekleştirilebilirler. Doğal olarak bazı zigotların ölümü bölünmenin doğasını etkiler.
Baskın genlerin resesif öldürücü etkilerle bölünmesine etkisi
Resesif öldürücü etkileri olan baskın genlerin pleiotropik etkisi vardır; bir yandan heterozigotta herhangi bir özelliğin tezahürünün baskın doğasını belirler, diğer yandan homozigottaki bireylerin ölümüne neden olurlar, yani öldürücü etkilerini resesif olarak gösterirler. Bu tür genler birçok hayvanda bilinmektedir; farelerde sarı renk, tavuklarda kısa bacaklar, sazanlarda doğrusal pullanma, tilkilerde platin rengi, koyunlarda şiraz rengi ve daha birçokları. 3:1 bölünmeden saparak 2:1 bölünmeye neden olurlar. Resesif öldürücü etkileri olan baskın genleri içeren ayrışmaların analizi, şu gerçek nedeniyle karmaşık hale gelir: nispeten küçük örneklerde 3:1 ve 2:1 bölünmeleri birbirinden ayırmak her zaman mümkün değildir ve χ2 kriterini kullanan istatistiksel test yöntemi, hipotezlerden seçim yapılmasına izin vermez. Örneğin, sarı fareler birbirleriyle çaprazlandığında 65 sarı: 32 siyah şeklinde bir bölünme elde edilirse, %2 yöntemi iki hipotezi reddetmez: 1 - deneydeki bölünme 3:1'lik bir bölünmeye karşılık gelir, χ 2 = 3,29, p>0,05; 2 - deneydeki bölme 2:1 bölmeye karşılık gelir, χ 2 = 0,17, p>0,05.
Yalnızca büyük örneklerde 2:1 bölünme, 3:1 bölünmeden ayırt edilebilir.. Farklı araştırmacılar tarafından elde edilen farelerde sarı rengin kalıtımına ilişkin verileri özetlerken, sarı fareleri birbirleriyle geçerken bölünme 2386 sarıydı: 1235 siyah - ∑ = 3621, χ 2 = 0,96, H 0 - 2:1 ( H 0 - 3:1'de bu durumda χ 2 = 160!).
Analitik ve geri melezlemeler gibi farklı genetik test yöntemlerinin kalıtım sorununun doğru çözümüne katkıda bulunabileceği açıktır. Yukarıda tartışılan örnekte bunlar haçlardır: F 1 × siyahtan sarı; F 1 × sarı P'den sarı. İlk durumda 1:1, ikincisinde ise yine 2:1 oranında bir bölünme olmalıdır.
Bazı zigotların ölümü postembriyonik aşamada meydana geldiğinde, bir sapma olduğunu ve bunun zigotların farklı yaşayabilirliğinden kaynaklandığını bulmak daha zordur. Bu bakımdan öldürücülük varsayımı varsa yavrularda doğurganlık ve ölüm oranının izlenmesi gerekir. Ölümcüllük varsayımının temeli, belirli bir fenotipteki bireyleri yetiştirirken tek tip yavrular elde etmenin imkansızlığıdır. Analiz örnekleri III.2 ve III.3 numaralı problemlerde verilmiştir.
Sorun No. III.2
F 1'de gümüş samur vizonların birbirleriyle melezlenmesinin yavrularında her zaman bölünme gözlenir: gümüş samur ve kahverengi bireyler ortaya çıkar.
Bir deneyde, birkaç gümüş-samur melezlemesinden 93 gümüş-samur ve 43 kahverengi yavru elde edildi ve ortalama yavru büyüklüğü 3,66 yavruydu. Gümüş samur ile kahverengi vizonların melezlenmesiyle 39 adet kahverengi ve 41 adet gümüş samur elde edilmiştir. Kahverengi vizonların birbirleriyle melezlenmesinden sadece kahverengi I yavruları elde edildi. Son iki melezlemedeki çöp boyutu 4,9-5,2 yavruydu.
Bölünmeyi açıklar, gümüş-samur ve kahverengi vizonların genotiplerini belirler.
Analiz
Gümüş samur vizonlar birbirleriyle ve kahverengi vizonlarla çaprazlandıklarında her zaman segregasyon ürettiklerinden, açıkça heterozigotturlar. Gen sayısını belirlemek için deneydeki bölünmeleri göz önünde bulundurun:
H 0 - bir gendeki farklılıklar, 3:1 bölünme, χ 2 = 3,2, p>0,05. Sapma rastgeledir, hipotez reddedilmez.

Oran, bir gen için 1:1 bölünmeye iyi bir şekilde karşılık gelir, χ2 = 0,05, p>0,80. Sapma rastgeledir, hipotez reddedilmez.
Farklı melezlerdeki vizonların doğurganlığına ilişkin verilerin karşılaştırılması, gümüş samur vizonları geçerken zigotların kısmi ölümünü göstermektedir. Baskın homozigotların öldüğünü varsayabiliriz. O zaman gümüş-samur vizonların genotipi Aa, kahverengi vizonlar - aa'dır ve melezlemedeki bölünme 3:1 değil 2:1'dir (χ 2 = 0,17, p>0,05). Bu hipotezin nihai olarak doğrulanması için gümüş samur bireyler arasında ek bir çaprazlama yapılması ve örneklemin arttırılması ve 2:1 hipotezinin istatistiksel olarak test edilmesi gerekmektedir. Bazı durumlarda daha karmaşık analizlerin yapılması gerekebilir.
Sorun No. III.3
Platin, beyaz yüzlü ve Gürcü beyaz tilkilerinin gümüş-siyah tilkilerle melezlenmesinde, pigmentasyonun genel olarak zayıflamasına ve çeşitli alacalı görünümlerin ortaya çıkmasına neden olan platin, beyaz yüzlü ve Gürcü beyazı renklerinin cinsiyet olmadığı tespit edildi. -bağlantılı. Bu mutantların her birinin gümüş-siyahlarla çaprazlanması 2:1'lik bir ayrışmayla sonuçlandı; gümüş-siyah rengi resesifti. Sonuç olarak, her biri resesif öldürücü etkiye sahip bir baskın gen tarafından kontrol edilir. Bazı mutantların ölümü, çöpün büyüklüğüne ilişkin verilerle kanıtlanmaktadır: gümüş-siyahların çöp başına 4,5 yavru, beyaz yüzlü 3,5, platin ve Gürcü beyazı - 3,5'ten az.
Bu mutasyonların bir geni mi yoksa farklı genleri mi etkilediğini tespit etmek için çaprazlamalar yapıldı ve sonuçları aşağıda verildi. Mutasyonlar baskın olduğundan allellik testi bu durumda uygulanamaz:

Bölünmeyi açıklar, tüm formların genotipini belirler.
Analiz
1. 1 ve 2 numaralı geçişlerdeki bölünmeler 1:1:1:1 oranına karşılık gelir (χ 2 = 5,83, 1 numaralı geçiş için p>0,10, 2 numaralı geçiş için χ 2 = 0,55, p>0,90) . Çeşitli nedenlerin sonucu olabilirler.
1. Çaprazlama 1 ve 2'de 4 bölünme sınıfı ve 1:1:1:1 oranı olduğundan, incelenen formların, aşağıdakilere göre etkileşime giren resesif öldürücü etkiye sahip, bağımsız olarak kalıtılan iki dominant gende farklı olduğu varsayılabilir. tamamlayıcılık türüdür. Bu durumda genlerden biri iki farklı baskın alel ile temsil edilir.

2. 1:1:1:1 ayrımı, aralarında geçiş olmadığında bu iki genin yakın bağlantısının bir sonucu olabilir:

(Punnett kafeslerinde fenotip radikalleri verilmiştir; fenotipte etkilerini gösteren genler.)
3. Renk farklılıkları, resesif öldürücü etkiye sahip, bağımsız olarak kalıtılan üç dominant genden kaynaklanır. Birinci (a) ve ikinci (b) melezlemelerde iki farklı gende bölünme meydana gelir.

4. 1:1:1:1 ayrışma, etkileşim halindeki üç genin çaprazlama olmadan yakın bağlanmasından kaynaklanabilir.

* (Genlerin sırası farklı olabilir; keyfi olarak verilmiştir.)
5. 1:1:1:1 oranındaki ayrışma, üçü resesif öldürücü etkiye sahip baskın ve dördüncüsü resesif olan dört alel dizisiyle temsil edilen bir gendeki farklılıkların sonucu olabilir:

Bu hipotezler arasında seçim yapabilmek için beyazlar ile gümüş-siyahlar arasında bir çarpı işareti yapıldı.
Bir özellik iki veya üç gen tarafından kontrol ediliyorsa, bağımsız kalıtımlarıyla birlikte dört fenotipik sınıfın ortaya çıkması beklenebilir:

Platin veya Gürcü beyaz tilkilerinden elde edilen beyazların gümüş-siyah tilkilerle çaprazlanmasında da benzer bir sonuç elde edilmeli, ancak beyaz yüzlü yerine platin olanların ortaya çıkması gerekir ki bu, ilgili haçların yazılmasıyla kolayca doğrulanabilir.
Test çaprazlamasında elde edilen bölünme - beyaz yüzlü ve Gürcü beyaz tilkilerinin görünümü - ya iki (veya üç) yakından bağlantılı genin etkileşimi ile ya da üç alelin orijinal arasındaki monogenik farkla etkileşimi ile açıklanabilir. formlar.

Deneyde not edilen beyaz tilkilerin yaşayabilirliğindeki keskin düşüş, alelik mutasyonların etkisi lehine konuşuyor, çünkü bu durumda beyaz tilkiler, her ikisi de resesif öldürücü etkiye sahip olan bir genin iki baskın mutasyonunun genotip bileşikleridir.İki farklı gen etkileşime girdiğinde (tamamlayıcılık) canlılığın azalmasını beklemek zordur. Bu nedenle, tilkilerdeki beyaz, beyaz yüzlü, Gürcü beyazı, platin ve gümüş-siyah renklerinin, üçü resesif öldürücü etkiyle baskın olan bir genin bir dizi alel tarafından kontrol edildiği sonucuna vardılar. Tilki genotipleri: beyaz A 1 / A 2, A 1 / A 3, A 2 / A 3; beyaz yüzlü A 1/a; platin A 3/a; Gürcü beyazı A 2/a; gümüş-siyah a/a (Belyaev ve arkadaşlarına göre, 1973).
Şunu vurgulamak gerekir ki Orijinal formların heterozigot olması durumunda çoklu alelizm ile, bölünmedeki maksimum fenotipik sınıf sayısı 3 değil 4 olabilir, yukarıda açıklanan durumda olduğu gibi. Bir popülasyonda çoklu allelizme sahip olası genotiplerin sayısı birçok kez artar; 1 / 2n (n+1) formülüyle belirlenebilir; burada n, alel sayısıdır. Örneğin bir lokus için 7 alel varsa popülasyondaki olası genotiplerin sayısı 28: 1/2 × 7 × 8 = 28 olacaktır.
Resesif öldürücü mutasyonların ayrışma üzerindeki etkisi
Resesif otozomal ve cinsiyete bağlı uçma,Ölümcül olarak heterozigotların çaprazlarında homozigotların ölümüne neden olurlar, öldürücü ile bağlantılı genlerin bölünmesini etkileyebilirler.Bu durumda, bölünmedeki torunların oranı, incelenen gen ile öldürücü arasındaki mesafeye ve ayrıca heterozigotun tipine göre - cis- veya trans-pozisyondaki genler heterozigota dahil edilir. Ölümcülleri belirlemek için genellikle çeşitli test çaprazlamaları yapılır. Uçuş analizine bir örnek, III.4 numaralı problemdir.
Sorun No. III.4
İnversiyonlar içeren 100 numaralı hattan Drosophila hattında dişilerin yarısı gri, yarısı sarı vücut rengine sahipti ve sarı dişilerin kısır olduğu ortaya çıktı. Bu soyun tüm erkekleri sarı renkteydi. Dişilerin erkeklere oranı normalden farklıydı; daha çok 2♀♀:1♂♂ şeklinde bir bölünmeye benziyordu. 100 numaralı çizgide erkeklerin bulunmamasının muhtemelen X kromozomunda açıkça heterozigot olan gri dişilerin varlığından kaynaklandığı ileri sürülmüştür; çizgi gri ve sarı dişilere bölünmüştür. Bu çizginin genetik yapısını oluşturmak ve dişilerin X kromozomunda ölümcül varlığın varlığının varsayımını test etmek için, sonuçları aşağıda sunulan çaprazlamalar yapıldı.
Karşılıklı haçlar

Gri dişiler F 1'den bireysel olarak 100 numaralı hattan sarı erkeklerle geçti.

Analiz
Analize dayanarak tüm geçişlerin şemalarını yazacağız.

Önerilen hipotezler tüm sonuçları oldukça iyi açıklamaktadır. Ancak 100. satırdaki sarı dişilerin kısırlığının nedeni sorusu hala belirsizliğini koruyor.Okuyucuyu bu soru üzerinde düşünmeye ve bunu açıklamak için bazı hipotezler önermeye davet ediyoruz.
sen bitkiler Birçok resesif mutasyon, klorofil eksikliği veya yokluğu ile ilişkilidir, bu da ya bitkinin yaşayabilirliğinde bir azalmaya ya da gelişimin çeşitli aşamalarında ölüme yol açar. Bu, bölünmede sapmalara neden olur ve aynı zamanda bitki ölümü oranını ve özelliğin kalıtım yapısını belirlemek için yalnızca fidelerde değil, aynı zamanda gelişimin sonraki aşamalarında da bölünmenin dikkate alınmasını gerekli kılar. Dolayısıyla mısır gen açısından homozigottur wd (beyaz eksikliği) beyaz fideleri var (fidanlara bölünmüş 3/4 yeşil: 1/4 beyaz). Ancak 1-3 hafta sonra, tohumun besin rezervleri tükendikten sonra tüm beyaz bitkiler ölür ve bitki gelişiminin en ileri aşamalarında bölünme kaybolur - 3 yeşil: 0 beyaz. Bezelye, arpa, çavdar, buğday vb.'de de benzer mutasyonlar bilinmektedir.
Diğer mutasyonlar, belirli bir gelişim aşamasındaki bireylerin yalnızca bir kısmının ölümüne neden olur, bu da bölünmedeki resesif oranında bir azalmaya ve fenotip oranında bir değişikliğe (4:1, 5:1 vb.) yol açar. Kural olarak bu tür mutantların yaşayabilirliği büyük ölçüde koşullara bağlı olduğundan bu oranlar değişiklik gösterir.
İnsanlarda Resesif mutasyonların etkisine bağlı olarak azalan canlılık ve öldürücülük, embriyogenezin farklı dönemlerinde ve gelişimin farklı aşamalarında kendini gösterir. Canlılıktaki azalma ve öldürücü etkinin nedenleri hem gen mutasyonları hem de kromozomal anormallikler ile ilişkilendirilebilir. Kürtajla alınan embriyoların sitogenetik analizi, birçoğunun ölüm nedenini belirlememize olanak sağlar. Ortalama olarak, gebeliğin tüm aşamalarında kromozom anormallikleri nedeniyle spontan düşüklerin %42'sinden fazlası meydana gelir: kromozomal anormallikleri olan yenidoğanların önemli bir kısmı yaşamın ilk ve sonraki yıllarında ölür.
Fetal ölüme veya bebeklik döneminde ölüme yol açan ölümcül gen mutasyonları arasında talasemi, orak hücreli anemi, kistik fibroz, konjenital iktiyoz, anensefali (beynin yokluğu), fenilketonüri vb. neden olan resesif mutasyonlar sayılabilir.
İnsanlarda öldürücü veya canlılığı azaltan mutasyonları incelemek için sitogenetik ve biyokimyasal analiz yöntemleri, sağlıkta ve hastalıkta ve ayrıca heterozigot taşıyıcılarda enzimlerin yapısını ve aktivitesini incelemek yaygın olarak kullanılmaktadır; kromatografi, farklı elektroforez türleri.
Ölümcül genler, bir bireyin cinsel olgunluğa ulaşmadan ölümüne neden olan mutasyonel genlerdir. Baskın, resesif veya cinsiyete bağlı olabilirler. Genellikle etkilerini homozigot durumda gösterirler; heterozigot durumda canlılığı azaltırlar. Penetrans, bir genin kendini fenotipik olarak gösterme yeteneğidir, % olarak ifade edilir ve tam veya eksik olabilir. Tam: Belirli bir gene sahip olan bir popülasyonun tüm bireylerinde bu, bir özellik olarak kendini gösterir. Eksik - bazı bireylerde gene sahiptir ancak kendisini dışarıdan göstermez. İfade gücü, bir özelliğin tezahür derecesidir, yani. Aynı özellik farklı bireylerde farklı yoğunlukta ifade edilir.
Farklı öldürücü genlerin varlığında organizmalar farklı gelişim aşamalarında ölürler. Kural olarak, bu tür genlerin öldürücü etkisi resesiftir, yani. yalnızca homozigot durumda olduklarında kendini gösterir. Baskın öldürücü etkiye sahip mutasyonlar meydana geldiğinde organizma yavru üretemeden ölür.
Bununla birlikte, heterozigot durumda gözle görülür değişikliklere neden olan öldürücü genlerin ekonomik açıdan yararlı hale geldiği durumlar da olmuştur. Böylece, Karakul cinsi koyunlar arasında, her zamanki siyah Karakul cinsine göre daha pahalı olan, güzel gümüş-gri ten rengine sahip hayvanlar vardır.Gri koyunları gri koçlarla geçerken, bunların her zaman heterozigot oldukları ortaya çıktı. Bu fenomenin nedenleri araştırılırken, böyle bir geçiş sonucunda elde edilen gri kuzular arasında toplam yavruların yaklaşık 1/3'ünün veya yaklaşık% 25'inin kronik timpanite yakalandığı ve öldüğü tespit edildi. Hastalığın nedeni parasempatik sinir sisteminin aktivitesindeki bozukluklardı. Gri koçlar kara koyunlarla veya gri koyunlar siyah koçlarla çaprazlandığında yavrular %50 gri ve %50 siyah kuzular üretti ve gri kuzular hastalanmadı. Homozigot durumda, gri rengin gelişmesine neden olan genin resesif, öldürücü bir etkiye sahip olduğu ortaya çıktı.
Tilkiler, homozigot durumda gelişimin erken bir aşamasında embriyoların ölümüne neden olan baskın bir genin neden olduğu değerli bir kürk rengine sahiptir - platin. Bu gibi durumlarda, ikinci nesildeki fenotiplerin oranı değişir, çünkü üç baskın formdan biri (ölümcül gen için homozigot) ölür ve bunun sonucunda fenotiplerin oranı 2: 1 olur.
Benzer bir bölünme, pulların azgelişmiş kalıtımının gözlendiği ayna sazanında da kaydedildi: yalnızca vücudun orta çizgisinde korunmuş, geri kalanı çıplaktı, bu yüzden doğrusal olarak adlandırılıyor. Lineer sazanlar birbirleriyle çaprazlandığında yavrular her zaman 2 lineer: 1 oranında normal pullarla bölünür, çünkü pulların az gelişmesine neden olan gen için homozigot olan embriyolar gelişimin erken aşamalarında ölür.
Tüy kıvrılmasına neden olan alel için homozigot olan tavuklarda, eksik tüy gelişiminin çeşitli fenotipik etkileri vardır. Bu tavukların ısı yalıtımı yetersiz olduğundan üşüme sorunu yaşıyorlar. Isı kaybını telafi etmek için bir takım yapısal ve fizyolojik adaptasyonlar geliştirirler, ancak bu adaptasyonlar etkisizdir ve bu tür tavuklar arasında ölüm oranı yüksektir.
Öldürücü bir genin etkisi, farelerde kürk renginin kalıtımında açıkça görülmektedir. Yabani farelerin genellikle agouti gibi gri kürkleri vardır; ancak bazı farelerin sarı kürkü vardır. Sarı fareler arasındaki melezlemeler 2:1 oranında hem sarı fareleri hem de agoutiyi üretir. Bu sonuçların tek olası açıklaması agutilerde sarı kaplama renginin baskın olması ve tüm sarı farelerin heterozigot olmasıdır. Atipik Mendel ilişkisi, homozigot sarı farelerin doğumdan önce ölümüyle açıklanmaktadır. Sarı farelerle çaprazlanan hamile sarı fareler üzerinde yapılan otopsilerde rahimlerinde ölü sarı yavrular ortaya çıktı. Eğer sarı fareler ve agotiler çaprazlanırsa, hamile kadınların rahimlerinde ölü sarı fareler kalmazdı, çünkü böyle bir melezlemeyle sarı yün geni için homozigot yavrular olamaz.
Monohibrit çaprazın ikinci neslinde fenotipik ayrışmada 3:1 oranındaki bir değişiklik, F2 zigotlarının farklı canlılığıyla ilişkilidir. Zigotların farklı yaşayabilirliği öldürücü genlerin varlığına bağlı olabilir. Ölümcül bir gen, bir organizmanın gelişiminde bozukluklara neden olan, onun ölümüne veya deformasyonuna yol açan bir gendir.
Konjenital anomalilerin incelenmesi, farklı öldürücü genlere sahip bireylerin ölümünün farklı olduğunu ve gelişimin farklı aşamalarında meydana gelebileceğini göstermiştir.
Rosenbauer (1969) tarafından önerilen sınıflandırmaya göre bireylerin %100'ünün olgunluğa ulaşmadan ölümüne neden olan genler öldürücü, %50'den fazlası sublethal (yarı öldürücü) ve %50'den azı subvital olarak adlandırılmaktadır. Ancak bu ayrımın bir dereceye kadar keyfi olduğunu ve bazen net sınırlarının bulunmadığını da belirtmek gerekir. Tavuklarda cinsiyete bağlı çıplaklık buna bir örnektir. Çıplak civcivlerin neredeyse yarısı kuluçka döneminin son 2-3 gününde ölür. Yumurtadan çıkan civcivlerin yaklaşık yarısı, 32-35 °C sıcaklıkta büyütülürse 6 haftalıktan önce ölür. Ancak kuluçka makinesindeki sıcaklık 5,5 °C artırılırsa, önemli ölçüde daha az çıplak civciv ölür. 4-- 5 ay sonra çıplak civcivlerin tüyleri seyrekleşir ve zaten oldukça düşük sıcaklıklara dayanabilirler.
Doğal koşullar altında bu mutasyonun öldürücü olması ve kuşların %100 ölmesine yol açması muhtemeldir. Yukarıdaki örnek, yarı öldürücü bir genin tezahürünün doğasının büyük ölçüde çevresel koşullara bağlı olabileceğini göstermektedir.
Ölümcül genler baskın veya resesif olabilir. İlk öldürücü faktörler arasında farelerin sarı rengine neden olan alel keşfedildi. Sarı renk geni baskındır (Y). Homozigot durumdaki resesif aleli (y), siyah rengin ortaya çıkmasına neden olur. Sarı farelerin birbirleriyle çaprazlanması, iki parça sarı fare ve bir parça siyah fare üretti; yani sonuç, Mendel kuralına göre 3:1 değil, 2:1 bölünme oldu. Tüm yetişkin farelerin heterozigot (Yy) olduğu ortaya çıktı. Birbirleriyle çaprazlandıklarında sarı renk (IT) için homozigot yavruların bir kısmını üretmeleri gerekirdi, ancak embriyonik dönemde ölürler, heterozigotların (Yy) iki kısmı sarı ve homozigotların bir kısmı sarı olacaktır. resesif özellik (yy) siyah olacaktır. Geçiş şeması şöyle görünür:
Aynı şekilde Karakul koyunlarında (Sokolskie, Malich vb.) gri kürk rengi, tilkilerde platin rengi, lineer sazanda pul dağılımı vb. kalıtsaldır.
Öldürücü genler çoğu durumda resesiftir ve bu nedenle uzun süre gizli kalabilirler.
Fenotipi tamamen sağlıklı ve normal olan bir hayvan, etkisi ancak homozigot duruma geçişte tespit edilen öldürücü bir genin taşıyıcısı olabilir. Ölümcül genler çoğunlukla akrabalı yetiştirme sırasında homozigot bir duruma geçer. At yetiştirirken hayvancılık uygulamasında, doğumdan sonraki 2.-4. günde 25 tayın rektal deformite - anüs yokluğu (Atresia ani) nedeniyle öldüğü bir vaka vardı. Bu tür anormal tayları doğuran tüm aygır ve kısrakların aynı aygırın soyundan geldiği ortaya çıktı. Ölümcül gen (LI) açısından heterozigottu. Başlangıçta bu aygır normal kısraklarla (LL) çaprazlandığında fenotipi normal olan yavrular doğurdu, ancak genotip açısından yavruların yarısı normal (LL) ve yarısı heterozigottu (LI), resesif eğilim (0 öldürücü gen. Heterozigot hayvanların (Y x Y) akrabalı çiftleştirilmesinde, ölümcül gen (II) için homozigot olan, rektal deformiteli bazı taylar ortaya çıktı. Hepsi öldü.
Çiftlik hayvanlarında, resesif veya dominant gen mutasyonlarıyla ilişkili olan düzinelerce anomali bilinmektedir. Bu anomaliler, mutasyon sürecinin hızına, hayvan yetiştirme sistemine vb. bağlı olarak farklı frekanslara sahip bireysel popülasyonlarda meydana gelir. Her türün hayvanlarındaki spesifik konjenital anomali formlarının yanı sıra bunların tezahürlerinin sıklığı hakkında bilgi genetik patolojinin yayılmasının seçici olarak önlenmesi için veteriner uzmanları için bireysel ırklar gereklidir.
Sığırlarda anomaliler. Bu tür hayvanların biyolojik özellikleri kısa tüylü olmaları ve nispeten geç olgunlaşmalarıdır. Bir inek genellikle bir buzağı doğurur ve bu buzağı cinsel ve fizyolojik olgunluğa ancak 1,5 yılda ulaşır, yani anne ile kızın ilk buzağılaması arasındaki süre ortalama 5 yıldır. Sonuç olarak, bir sürüde anormal yavruların ortaya çıkması, üreme düzeyini ve çiftlik hayvanlarının üreme seçiminin yoğunluğunu önemli ölçüde azaltabilir. Sığırlarda öldürücü, yarı öldürücü ve subvital genler tarafından belirlenen çok çeşitli konjenital shamalia araştırılmıştır. Uluslararası Ölümcül Kusurlar Listesi'nde A kodu altında 46 anormallik yer almaktadır (Tablo 43). Her cins veya popülasyondaki belirli anormallik türlerinin göreceli sıklığı değişebilir. Verilerimize göre Kostroma ırkında en sık kaydedilen genetik kafa anomalisi çene kısalmasıdır (Tablo 44), Yaroslavl ırkında sindaktili, Kholmogory ırkında kas kontraktürleri, Black-and- Beyaz - göbek fıtığı. Almanya'daki sığırlarda merkezi sinir sistemi anormallikleri en yaygın olanıydı (%21).
Kayıt sıklığında ikinci sırada (% 14), karmaşık bir anomali - göbek fıtığı ile yarık karın ve bir bütün olarak fetüsün birleşimi - işgal edildi. Belirli popülasyonlardaki anormalliklerin sıklığı veya anormal yavruların toplam sayıya oranı da çok farklı olabilir ve ortalama tahminlere göre %1'i aşmaz. Ancak bu gösterge, anormallikleri kaydetmenin eksiksizliğine ve doğruluğuna bağlıdır. Böylece Almanya'da net bir muhasebe düzenlendikten sonra anormallik sıklığının birkaç kat arttığı sonucuna vardılar. Soru şudur: Tüm anormallikler görsel gözleme uygun mudur? Açıkçası hepsi değil. Böylece Kostroma ırkında 12 yıllık bir süre boyunca tüm anomali türlerinin ortalama sıklığı %1,15 oldu. Bu çiftlikte genel çöp ölüm oranı (düşük, ölü doğmuş, anormal, görünür kusurları olmayan ölü buzağılar) %10,2 idi. Bu ölümlerin belli bir kısmı, morfolojik bozukluklara değil, tanımlanması ancak özel yöntemlerle mümkün olan metabolik bozukluklara ve diğer anormalliklere neden olan gen mutasyonlarıyla da ilişkilidir.
Üreticiler hem sığırlarda hem de diğer hayvan türlerinde genetik anormalliklerin yayılmasında özel bir rol oynayabilir. Her bir babadan suni tohumlama ile yılda yüzlerce, binlerce yavru elde edilebilmektedir. Böylece yurtdışındaki 1 boğadan 100 bin buzağı elde edildi. Eğer böyle bir babanın bir gen mutasyonunun taşıyıcısı olduğu ortaya çıkarsa, bu durum hızla tüm ırka yayılacaktır. Literatürde açıklanan çok sayıda olgudan bazı örnekleri burada bulabilirsiniz. İsveç'e getirilen Prens Adolf boğasının yoğun kullanımı ve daha sonra kendiliğinden akraba çiftleşmesi sonucunda bazı İsveç sürülerinde tüysüzlük oranı %5'in üzerinde çıkmıştır. Aynı durum, uzuvların yokluğuna neden olan genin heterozigot bir taşıyıcısı olduğu ortaya çıkan Gallus boğasının ithal edilmesinden sonra İsveç'te de ortaya çıktı.
ABD ve Almanya'da bireysel Siyah-Beyaz ve Charolais boğalarının yavrularında sırasıyla %23,3 ve %22,2 sıklıkta cüce buzağı doğum vakaları kaydedildi. Eskiden Çekoslovakya'da 166 babanın torunları incelenirken bunlardan 43'ünün öldürücü gen taşıyıcısı olduğu ortaya çıktı. Baskın anomali olan "yarık dudak"ın taşıyıcısı olan bir boğada, kusur, boğaların %44'ünde ve onun yavrularından elde edilen düvelerin %71'inde kendini gösterdi.
Kostroma ırkında alt çene kısalığı ve boksör benzeri görünümün yayılımını, kendi yavrularında kusurlu buzağılara sahip olan Burkhan boğası üzerinden; oğulları, torunları, torunlarının çocukları, kadın torunları da anormal yavrular verdi (Şekil 57). Buzağıların çoğu, normal fenotipe ve ortak ataya sahip ebeveynlerin akraba evliliğinden ve çiftleştirilmesinden elde edilir. Dolayısıyla bu anomalinin resesif kalıtım moduna sahip olduğu sonucuna varabiliriz. Şekil, en fazla sayıda kusurlu buzağının, bazı ineklerin genotiplerinde aynı resesif gene sahip olduğu ticari bir çiftlikte kullanıldığında boğa Zheton 3501'in (Burkhan boğasının torunu) yavrularında kayıtlı olduğunu göstermektedir.
Domuzlarda anomaliler. Domuzlardaki Ölümcül Kusurların Uluslararası Listesi 18 genetik anormallik içermektedir. Bunların büyük bir kısmı otozomal resesif genlerden kaynaklanmaktadır (Tablo 45). Genetik anormallikler domuzların patolojisinde önemli bir rol oynayabilir. Bu tür birkaç örneğe bakalım. İspanya'da Duroc, Yorkshire, Hampshire ve White Chester cinsi domuzlardan elde edilen 2.399 yavrudan 23.449 domuz yavrusu üzerinde yapılan bir araştırma sırasıyla 6.21; 6.02; 9.66; Anormal yavruların %2^62'si.
Olivier (1979)'e göre 7'si deride, 17'si iskelette, 3'ü gözlerde, 13'ü nöromüsküler, 6'sı kanda, 6'sı hormonal metabolizmada, 5'i sindirim sisteminde ve 9'u Domuzlarda genitoüriner sistem tanımlanmıştır. Başlıca anomaliler kriptorşidizm, fıtıklar, psödohermafroditizm vb. idi. Araştırmanın yazarı, bu anormalliklerin embriyo oluşumunun farklı aşamalarında bir genin etkisinin sonucu olduğuna inanmaktadır.
Danimarka'da, konjenital anomalilerin doğasını ve sıklığını belirlemek için iki yıllık bir süre boyunca 2.936 yavrudan 6.669 ölü domuz yavrusu üzerinde çalışıldı. Doğan domuz yavrularının yüzde 1,4'ünde, sütten kesilmeden öldürülenlerin ise yüzde 6,2'sinde çeşitli anomaliler tespit edildi. Otopsi sırasında, anormal domuz yavrularının %25,9'unda az gelişmiş kapakçıklar, anüsün kapanmaması, subaortik stenoz, kalpte ektopi ve kardiyovasküler sistemdeki diğer kusurlar olduğu tespit edildi. Domuz yavrularının %23,4'ünde motor sistemin çeşitli gelişimsel bozuklukları tespit edildi. Domuz yavrularının %5,9'unda, çatallanmış beyin ve serebral hidrosel de dahil olmak üzere merkezi sinir sistemi anomalileri tespit edildi. Domuz yavrularının %30'unda rektumun aşırı büyümesi, ince bağırsaklar veya bunların eksik gelişimi ve %6,8'inde çeşitli fıtıklar ve asitler bulundu. “Yarık dudak”, yarık damak, rinosefalit ve başın yüz kısmındaki diğer anomaliler %6,1; hermafroditizm, yarık üreter, böbreklerde ve üretrada hidrosel - domuz yavrularının %1,7'sinde. Bu anomaliler, akrabalı yetiştirme sırasında bireysel babaların yavrularında ortaya çıktı; bu da, bunların oluşumunun kalıtsal doğasını gösteriyor.
Domuzlarda kriptorşidizmin kalıtsal doğasına dair çok ikna edici kanıtlar Fridin ve Newman tarafından elde edildi. Verilerine göre Kanada'da her yıl piyasaya giren tüm domuzların %1-2'sinde tek taraflı ve iki taraflı kriptorşidizm görülüyor. Yazarlar kriptorşidleri anneleri ve öz kız kardeşleriyle çaprazladılar. Bu tür melezlemelerden elde edilen yavrular birbirleriyle çiftleştirildi. Bu tür bir seleksiyon ve seleksiyon sonucunda Yorkshire cinsi deney hayvanlarında ve özellikle iki üretici kullanıldığında kriptorşidizm görülme sıklığı ortalama %42,9'a çıkmıştır. Amerika Birleşik Devletleri'ndeki domuz yavrularını bir yılda incelerken, skrotal fıtıklı yaklaşık 400 bin hayvan bulundu.
Gözlemler, domuzlarda doğurganlığın bozulmasının nedeninin sıklıkla testislerin hipoplazisi olduğunu göstermektedir. Almanya'dan araştırmacılara göre bu anomalinin görülme sıklığı %19,6 idi. Bu tür 30 domuz üreme için bırakıldı
Va, her biri 4 ila 40 kraliçe (toplam 439 baş) kapsıyordu, ancak bunlardan yalnızca dördü yavru doğurdu. Analiz, bu domuzlardaki spermin patolojik formlarının% 80-100 olduğunu gösterdi. 30 anormal hayvanın hepsinin ortak ataları vardı, bu da testis hipoplazisinin ve spermiyogenez kusurlarının kalıtsal doğasını gösteriyor.
Domuzlarda krater meme uçlarının varlığı ciddi kusurlardan biridir çünkü domuz yavruları onlardan süt almaz. Bavyera Hayvancılık Enstitüsü'ne (Almanya) göre, Alman Yerel Çeşitlerinde bu anomalinin sıklığı %6,6 idi. P. N. Kudryavtsev ve arkadaşlarının (MVA) belirttiği gibi, aktif olmayan krater memelerine sahip domuzların sayısı son yıllarda arttı. Bu tür meme uçlarının sayısı 1 ila 8 arasında değişmektedir. Krater meme uçlarını alan domuz yavruları ölür.
Kraterlik, otozomal resesif bir genin neden olduğu bir özelliktir. Bu, P. N. Kudryavtsev ve diğerleri tarafından deneysel olarak test edilmiştir. Anomalilerin (KchKch) taşıyıcısı olan domuzları ve yaldızları, yavrularında kraterli domuz yavruları üreten normal fakat heterozigot bireyleri (KchKch) ve normal homozigot domuzları (KhKch) daha önce tanımlamış olan yazarlar, bu hayvan grupları arasında melezlemeler gerçekleştirdiler. . İlk varyantta, 27 normal homozigot baraj 15 domuzla çaprazlandı. 258 çocuğun tamamı normaldi. Ebeveynlerden birinin homozigot (KchKch), diğerinin ise heterozigot (Kchkch) olduğu ikinci seçenekte ise tüm domuz yavruları da normaldi. Üçüncü varyantta 13 heterozigot domuz, 16 heterozigot anne ile çaprazlandı. Doğan 168 domuz yavrusundan 39'unda (%23,2) krater meme uçları vardı. Ve son olarak dördüncü seçenekte ebeveynlerden biri homozigot, diğeri ise heterozigottu. 170 domuz yavrusu ürettiler; bunlardan 86'sı (%50,5) normal meme uçlarına, 84'ü (%49,5) ise krater meme uçlarına sahipti. Bu deneyin sonuçları domuzlarda meme başı kraterinin resesif kalıtım tarzını kanıtlıyor.
Koyunlarda anormallikler. Koyunlarda yaklaşık 90 kadar konjenital anomali tanımlanmıştır. Dennis ve Leipold'a göre koyunlarda bilinen genetik kusurların çoğu monogenik otozomal resesif kalıtım tarzından kaynaklanmaktadır (Tablo 46). Bu hayvan türlerinde en yaygın kraniyofasiyal kusurlar, kraniyofasiyal kusurlar, özellikle agnati, ayrıca ön ayakların eğriliği, mikroagnati, hermafroditizm, kriptorşidizm, hipospadiazis, prognati, anal atrezi, mikrotia, entropi, tortikollis, polythelia, artrogripozdur. Analiz, kusurların %55,4'ünün kas-iskelet sistemi, 12,7'sinin sindirim sistemi, 9,7'sinin kardiyovasküler sistem, 7,1'inin ürogenital sistem, 6'sının merkezi sinir sistemi, 3,5'inin anomali ile ilgili olduğunu gösterdi. bağların 3,2'si karın bölgesine, %1,5'i endokrin sisteme. Bireysel kusurların sıklığı düşük olmasına rağmen tüm anormalliklerin kümülatif katkısı çiftliklere zarar verebilmektedir. Koyun yetiştiriciliğinin gelişmiş olduğu bir ülke olan Yeni Zelanda'da, ölümcül kusurların görülme oranı ölü kuzuların yaklaşık %1'i kadardı. ABD'de kuzuların %11,4'ünde yaşamın ilk 2 haftasında ölümcül kusurlar gözlendi.
Koyunlarda ortalama embriyo ölüm oranı %20'dir. Bu durum, tanımlanamayan birçok öldürücü genin bu dönemde aktif olabileceğini göstermektedir.
Koyunlarda bireysel genetik anormallikler yaygın olabilir. Böylece Bulgaristan'da Merinos koyunu sürülerinde doğum sonrası erken dönemde yüksek kuzu ölümleri gözlendi. Kuzuların, annelerinin memelerindeki kusurlar nedeniyle süt alamamaları sonucu ortaya çıktı: küçük glandüler doku kalıntıları olan hipoplaziden tamamen yokluğuna kadar. Bu kalıtsal anomalinin farklı sürülerdeki sıklığı %6 ile %40 arasında değişmektedir.
Koyunlarda, yoklama gibi ikincil bir cinsel özellik ile birleştirilen kriptorşidizm sıklıkla gözlendi. Boynuzsuz koçların doğurganlığı düşüktür. Sıkı üreme seçimi yoluyla, normal doğurganlığa sahip bir tür boynuzsuz koç yaratmak mümkün olmuştur, ancak bunların arasında anormal bireyler de bulunur.
Kuşlarda anomaliler. Anormalliklerin genetiği açısından en çok araştırılanlar kuşlar, özellikle de tavuklardır. Uluslararası Ölümcül Kusurlar Listesi tavuklarda 45, hindilerde 6 ve ördeklerde 3 anomali içermektedir. En sık görülen gaga anormallikleri (papağan gagası, çapraz gaga) şeklindedir. William ve arkadaşlarına göre bunların sıklığı, White Leghorn ve Rhode Island tavuklarından elde edilen yumurtaların kuluçkalanması sırasındaki kayıpların %1,1'idir. Ördeklerde gaga anormallikleri de yaygındır.
Max Gibbon ve Shakelfurd, beyaz leghornları tereyağlılar ve ufak tefeklerle geçerken ve ardından "kendi içinde" üreme - polidaktili - sırasında bir anormallik tanımladılar. Ayrıca tavuklarda sindaktili ve tüylü bacaklar gözlendi. Sendromun görülme sıklığı %16,8 idi. Anormal bir F2 horozu fenotipik olarak normal tavuklarla geçerken, yarı normal ve yarı anormal bireylerden oluşan bir bölünme gözlendi (1:1). Anormal tavukların embriyonik ve embriyo sonrası hayatta kalma oranı çok düşük olduğundan, bu özellik kompleksinin yarı öldürücü etkiye sahip bir otozomal gen tarafından kontrol edildiği tespit edilmiştir.
Atlarda anomaliler. Atlardaki kalıtsal anomalilerden 10'u Uluslararası Ölümcül Kusurlar Listesi'nde yer almaktadır. Bunlar arasında 3'ü iskelet, 2'si üreme sistemi, 2'si böbrek ve kas anomalisi, 1'er bağırsak, sinir sistemi ve görme organı anomalisi yer alıyor.
Taslak ırk atlarda kolon atrezisi daha yaygındır. Bu anomalinin yaygınlığı Percheron cinsi Superba aygırının yavrularında görülmüştür. Aynı zamanda Doğu Afrika kökenli safkan bir binici ırkının taylarında da tanımlanmıştır. Taslak ırklarda kusurlu epiteliyogenez kaydedilmiştir. Oldenburg adı verilen ataksi, Almanya'daki Oldenburg taylarında bulundu. 9. hatta yayıldı. Atlarda en sık kaydedilen anomalilerden biri göbek fıtığıdır. Hafif ve ağır ırklarda bulunur.
Atlarda başka birçok genetik ve kalıtsal-çevresel anomaliler de bilinmektedir. Bu nedenle, ABD'de çeşitli cinslerdeki atların "overo" adı verilen tuhaf beyaz benekli tayları vardır. "Overo" tipi atları geçerken, pembe tenli taylar doğar; burada bağırsak hipoplazisi ve izoeritrolizin yanı sıra kolik ölüme yol açar.
İngiltere'de, hareket koordinasyonunda bozulma olan hayvanlar - "yalpalama hastalığı" - melez binicilik cinsinin atlarında kaydedildi ve incelendi. Bu anomaliye genetik bir yatkınlık kurulmuştur.
Atlarda ekstremite dermatozlarının kalıtsallığı kanıtlanmıştır. Kalıtsal yatkınlığa sahip kusurlar, diz ekleminin sıklıkla meydana gelen kronik deforme edici iltihabıdır - taylarda "koltuk değneği bacağı" olarak adlandırılan direk ve ayrıca esas olarak yarış atlarında ve yarış atlarında gözlenen toynakların koronal bloğunun kronik aseptik iltihabıdır. atlar.









- 01'den sonra edinilen taşınır mallar
- 01'den sonra edinilen mülklere ilişkin vergi
- Denizatı - mesaj raporu Denizatı dünyada ne kadar süredir var?
- Sütlü çikolatalı vanilyalı kekler
- Fotoğraflı ekşi mayalı krep tarifi
- Votkanın icadının tarihi
- Kutsal Babaların Mirası: Yaşam ve ahlakla ilgili Ortodoks benzetmeler ve İncil'den alıntılar
- Kraliyet eti, kraliyet eti pişirmek için Yeni Yıl tarifi Fırında patatesli kraliyet eti
- Okroshka için beyaz kvas Evde okroshka kvası yapın
- Kimya (Atom yapısı) (sunum)
- İngiltere'nin sizi şaşırtacak gelenek ve görenekleri
- Kalıtım ve gelişme Kalıtım bu kadar önemli mi?
- İngilizce Harry Potter kitapları
- Strese neden olan örgütsel faktörler
- Yavaş bir tencerede haşlanmış lahana tarifi: basit, hızlı ve lezzetli
- Değnek Sekizlisi, kartların tanımı ve özellikleri
- Biftek ve somon madalyonları
- Ses otomasyonuna ilişkin bireysel konuşma terapisi dersinin özeti Cümlelerde ses otomasyonunun özeti l
- Nizhny Novgorod Devlet Pedagoji Üniversitesi adını aldı
- İsa'yı çarmıha gerenlerin hepsi korkunç bir şekilde öldüler. Pontius Pilatus gerçekten orada mıydı?