Recesivni, dominantni, smrtonosni geni, mutacije. III.2.8
Do sada smo ljudsku genetiku razmatrali po vrstama nasljeđivanja, a sada ćemo se osvrnuti na jednu vrlo važnu kategoriju gena, a to su smrtonosni i subletalni geni. Letalni i subletalni geni su klasa mutacija sa izrazito izraženim negativnim efektima na razvoj. Kao što ćemo kasnije vidjeti, mutacije u mnogim slučajevima pogoršavaju i uništavaju naslijeđe osobe. Ovo


pogoršanje mutacijama posebno je izraženo pod uticajem smrtonosnih gena.
Letalni i subletalni aleli mogu biti dominantni ili recesivni. U slučaju dominantne letalnosti, dijete umire u prenatalnom periodu ili u ranom djetinjstvu. Sasvim je jasno da se takvi geni ne mogu naslijediti. Njihovo prisustvo ukazuje na pojavu mutacija. Ukupna učestalost takvih dominantnih smrtonosnih slučajeva nije tako mala. Njihov broj je posebno veliki pod uticajem zračenja. U genetici miševa i drozofile dominantni letalci se široko koriste kada se analizira zavisnost učestalosti mutacije o dozi i kada se analiziraju druge karakteristike mutagenih efekata. U prisustvu subletala, inače zvanih poluletalni geni, dijete umire u ranom djetinjstvu ili nešto kasnije, ali po pravilu prije puberteta. Primjer dominantne subletalne mutacije je epiloja, bolest koju karakterizira učinak cijelog sindroma negativnih simptoma. Djeca sa epiloijom imaju patološke izrasline na koži, karakteriziraju ih mentalna retardacija, epilepsija, tumori srca, bubrega i drugih organa. U rijetkim slučajevima, pacijenti s epiloijom prežive i stupe u brak sa zdravim supružnikom. Ispostavilo se da je polovina djece iz takvog braka bolesna od urođene epiloje (Sl. 131).
Još jedan upečatljiv primjer subletalne dominantne mutacije je bolest retinoblastoma. Ovo


dominantni gen izaziva kancerogeni tumor u oku, koji gotovo uvijek dovodi do smrti djeteta u ranoj dobi.
Međutim, najveći dio kongenitalnih deformiteta pada na recesivne letalne i subletalne. U ovom slučaju, recesivni štetni geni su skriveni u genotipu naizgled zdravih roditelja, međutim, kada se dva takva heterozigota vjenčaju, oko 25% njihove djece se ispostavi da je bolesno.
Na slici 132 prikazane su glavne vrste ukrštanja u ljudskim populacijama koje se vrše pri nasljeđivanju recesivnih smrtonosnih i subletalnih gena. Na kvadrat 1 predstavlja tip braka između genetski normalnih ljudi. Kod oba roditelja i svih njihovih potomaka, svi aleli su predstavljeni normalnim genima (alel +). Na kvadrat 2 prikazuje brak između genetski zdrave osobe i heterozigota (simbol a). U ovom slučaju, negativni alel se prenosi na polovinu potomstva, ali ostaje skriven. Na kvadrat 3 genetski zdrava osoba udaje se za bolesnu osobu (simbol aa), sva djeca su naizgled zdrava, ali u heterozigotnom stanju nose subletalnu mutaciju A. Na kvadrat 4 heterozigot se udaje za pacijenta; polovina njihove djece je naizgled zdrava, ali nosi subletalnu mutaciju, druga polovina je bolesna. Konačno, kvadrat 5 u slučaju braka dvoje bolesnih osoba (simboli aa) svo potomstvo će biti bolesno. Urođene bolesti uzrokovane djelovanjem recesivnih smrtonosnih i subletalnih gena uključuju dječju amaurotičnu idiotiju, u kojoj degenerira moždano tkivo, dolazi do sljepoće i dijete umire; urođena patologija kože s prisutnošću
duboke krvareće pukotine - ihtioza (Sl. 133); kongenitalna infantilna paraliza i niz drugih urođenih bolesti.
Mnogi rani spontani pobačaji uzrokovani su curenjem smrtonosnih genotipova.
Poznate su dominantne mutacije koje kod heterozigota određuju neke više ili manje neutralne osobine, koje, međutim, imaju smrtonosni učinak u homozigotnom stanju. Na primjer, alel pigmentacije kseroderme u heterozigotnom stanju uzrokuje jake pjege. Međutim, homozigoti razvijaju xeroderma pigmentosum. Ovo je subletalna bolest (Sl. 134), kod koje djelovanje svjetlosti uzrokuje oštećenje kože, što potom dovodi do malignog rasta.
Jedan od alela kod osobe u heterozigotnom stanju uzrokuje višak kolesterola u krvi. Međutim, djeca homozigotna za ovaj alel razvijaju tešku subletalnu abnormalnost.
U svim ovim slučajevima imamo klasičnu sliku nasljeđivanja dominanta s recesivnim letalnim efektom, prethodno utvrđenu u eksperimentima s drozofilom, miševima i drugim organizmima. Na primjer, miš ima dobro poznat alel A Y, što uzrokuje da koža postane žuta kod heterozigota. Međutim, svi pokušaji da se dobije čista rasa žutih miševa ostali su neuspješni. Prilikom ukrštanja dva žuta miša, u prosjeku, dva žuta miša uvijek proizvedu jednu druge boje, na primjer crnu. Priroda ovog fenomena objašnjena je činjenicom da se pokazao gen za dominantnu žutu boju
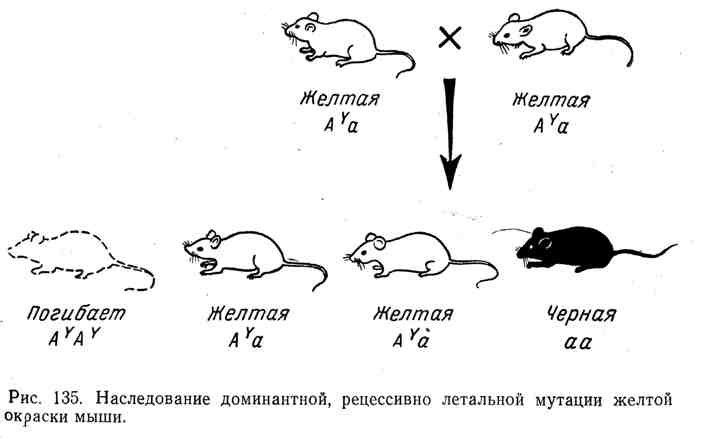
recesivno smrtonosno. Svi homozigoti( A Y A Y) umro. Žuti miš je uvijek heterozigotan, zajedno sa alelomA Y sadrži drugi alel, npr. A(crna boja). Tok nasljeđivanja pri ukrštanju heterozigota A Y a i cijepanje prema formuli 2:1 prikazani su na slici 135.
- Izvor-
Dubinin, N.P. Horizonti genetike / N.P. Dubinin. – M.: Prosveta, 1970.- 560 str.
Broj pregleda: 121
Nejednaka održivost zigota različitih genotipova može biti povezana s dominantnim ili recesivnim smrtonosnim mutacijama koje nastaju u gametama roditeljskih jedinki. Mogu se realizovati u različitim fazama embriogeneze ili u postembrionalnom periodu. Naravno, smrt nekih zigota utiče na prirodu cijepanja.
Utjecaj na cijepanje dominantnih gena sa recesivnim letalnim efektima
Dominantni geni sa recesivnim letalnim efektima imaju pleiotropni efekat; s jedne strane određuju dominantnu prirodu manifestacije bilo koje osobine kod heterozigota, s druge izazivaju smrt jedinki u homozigotu, odnosno ispoljavaju svoje smrtonosno djelovanje kao recesivno. Takvi geni su poznati kod mnogih životinja - žuta boja kod miševa, kratke noge kod pilića, linearna ljuska kod šarana, platinasta boja kod lisica, boja shirazi kod ovaca i mnogi drugi. Oni uzrokuju odstupanje od dekoltea 3:1, pretvarajući ga u dekolte 2:1. Analiza segregacija koje uključuju dominantne gene sa recesivnim letalnim efektima je komplikovana činjenicom da u relativno malim uzorcima, nije uvijek moguće razlikovati podjele 3:1 i 2:1, a statistička metoda testiranja pomoću χ 2 kriterijuma ne dozvoljava izbor između hipoteza. Na primjer, ako se pri ukrštanju žutih miševa dobije razdioba od 65 žutih: 32 crnih, metoda %2 ne odbacuje dvije hipoteze: 1 - podjela u eksperimentu odgovara razdvajanju od 3:1, χ 2 = 3,29, p>0,05; 2 - cijepanje u eksperimentu odgovara cijepanju 2:1, χ 2 = 0,17, p>0,05.
Samo u velikim uzorcima može se razlikovati 2:1 podjela od 3:1 podjela.. Kada se sumiraju podaci o nasljeđivanju žute boje kod miševa koje su dobili različiti istraživači, podjela pri ukrštanju žutih miševa međusobno je bila 2386 žutih: 1235 crnih - ∑ = 3621, χ 2 = 0,96 pri H 0 - 2:1 ( na H 0 - 3:1 u ovom slučaju χ 2 = 160!).
Očigledno je da različite metode genetskog testiranja, na primjer, provođenje analitičkih i povratnih ukrštanja, mogu doprinijeti ispravnom rješavanju pitanja nasljeđivanja. U primjeru o kojem se gore govori, ovo su križevi: žuti od F 1 × crni; žuti od F 1 × žuti P. U prvom slučaju bi trebalo biti podjela 1:1, u drugom - opet 2:1.
Teže je utvrditi da postoji odstupanje i to zbog različite vitalnosti zigota u slučaju kada do smrti nekih zigota dođe u postembrionalnoj fazi. S tim u vezi, potrebno je pratiti plodnost i mortalitet potomaka ako postoji pretpostavka o smrtnosti. Osnova za pretpostavku smrtnosti je nemogućnost dobivanja ujednačenog potomstva prilikom uzgoja jedinki određenog fenotipa. Primjeri analize dati su u zadacima br. III.2 i III.3.
Problem br. III.2
U potomstvu ukrštanja kune srebrnog sablea jedna s drugom u F 1, uvijek se uočava cijepanje: pojavljuju se jedinke srebrnog sable i smeđe.
U jednom eksperimentu, nekoliko legla ukrštanja srebrnog samura dalo je 93 šteneta srebrnog i 43 smeđa, sa prosječnom veličinom legla od 3,66 štenaca. Ukrštanjem srebrnog samura sa smeđim kunama dobijeno je 39 smeđih i 41 srebrni samur. Ukrštanjem smeđih minka međusobno su dobiveni samo smeđi I potomci. Veličina legla u zadnja dva ukrštanja bila je 4,9-5,2 štenaca.
Objasniti cijepanje, odrediti genotipove srebro-sable i smeđe kune.
Analiza
S obzirom na to da kune srebrnog sable uvijek proizvode segregaciju kada se ukrste jedna s drugom i sa smeđim, očito su heterozigotne. Da biste odredili broj gena, razmotrite podjele u eksperimentu:
H 0 - razlike u jednom genu, cijepanje 3:1, χ 2 = 3,2, p>0,05. Odstupanje je nasumično, hipoteza se ne odbacuje.

Odnos dobro odgovara podeli 1:1 za jedan gen, χ 2 = 0,05, p>0,80. Odstupanje je nasumično, hipoteza se ne odbacuje.
Poređenje podataka o plodnosti kune u različitim ukrštanjima ukazuje na djelomičnu smrt zigota pri ukrštanju kune srebrnog samura. Možemo pretpostaviti smrt dominantnih homozigota. Tada je genotip kune srebrnog samura Aa, smeđih - aa, a cijepanje u križanju nije 3:1, već 2:1 (χ 2 = 0,17, p>0,05). Za konačnu potvrdu ove hipoteze potrebno je izvršiti dodatno ukrštanje jedinki srebrnog samura kako bi se povećao uzorak i statistički testirala hipoteza 2:1. U nekim slučajevima potrebno je izvršiti složeniju analizu.
Problem br. III.3
U ukrštanju platinaste, bele i gruzijske bele lisice sa srebrno-crnim, utvrđeno je da platinasta, belolika i gruzijsko bijela boja, koje izazivaju opšte slabljenje pigmentacije i pojavu raznih pegastih lisica, nisu polne. -povezano. Ukrštanje svakog od ovih mutanata sa srebrno-crnim rezultiralo je segregacijom 2:1, pri čemu je srebrno-crna boja bila recesivna. Prema tome, svaki od njih je pod kontrolom jednog dominantnog gena sa recesivnim smrtonosnim efektom. O smrti nekih mutanata svjedoče podaci o veličini legla: srebrno-crni imaju 4,5 štenaca po leglu, bijelo lice 3,5, platinasto i gruzijsko bijelo - manje od 3,5.
Kako bi se utvrdilo da li su ove mutacije zahvatile jedan gen ili različite, izvršeno je ukrštanje čiji su rezultati dati u nastavku. Test za alelizam u ovom slučaju nije primjenjiv, jer su mutacije dominantne:

Objasniti cijepanje, odrediti genotip svih oblika.
Analiza
1. Podjele u ukrštanjima 1 i 2 odgovaraju omjeru 1:1:1:1 (χ 2 = 5,83, p>0,10 za ukrštanje 1, χ 2 = 0,55, p>0,90 za ukrštanje 2) . One mogu biti posljedica više razloga.
1. Kako u ukrštanjima 1 i 2 postoje 4 klase cijepanja i odnos 1:1:1:1, može se pretpostaviti da se proučavani oblici razlikuju po dva nezavisno naslijeđena dominantna gena sa recesivnim letalnim efektom koji međusobno djeluju prema tip komplementarnosti. U ovom slučaju, jedan od gena je predstavljen sa dva različita dominantna alela.

2. 1:1:1:1 segregacija može biti posljedica bliske veze ova dva gena u odsustvu križanja između njih:

(U Punnettovim rešetkama dati su fenotipski radikali - geni koji manifestiraju svoj učinak u fenotipu.)
3. Razlike u boji uzrokuju tri nezavisno naslijeđena dominantna gena sa recesivnim letalnim efektom. U prvom (a) i drugom (b) ukrštanju dolazi do cijepanja u dva različita gena.

4. Segregacija 1:1:1:1 može biti rezultat bliske veze tri gena u interakciji u odsustvu križanja.

* (Redoslijed gena može biti različit, dat je proizvoljno.)
5. Segregacija u omjeru 1:1:1:1 može biti rezultat razlika u jednom genu, predstavljenom nizom od četiri alela, od kojih su tri dominantna sa recesivnim letalnim efektom, a četvrti je recesivan:

Da bi se napravio izbor između ovih hipoteza, napravljen je ukrštanje - bijeli sa srebrno-crnim.
Ako bi osobinu kontrolisala dva ili tri gena, onda bi se uz njihovo nezavisno nasljeđivanje očekivala pojava četiri fenotipske klase:

Sličan rezultat bi trebao biti i kod ukrštanja bijelih lisica dobivenih od platinastih ili gruzijskih bijelih lisica sa srebrno-crnim, međutim, umjesto bijelih, trebale su se pojaviti platinaste, što se lako može provjeriti ispisivanjem odgovarajućih križeva.
Podjela dobivena u probnom ukrštanju - pojava bijele i gruzijske bijele lisice - može se objasniti ili interakcijom dva (ili tri) blisko povezana gena, ili interakcijom tri alela s monogenom razlikom između originalnih forme.

Naglo smanjenje vitalnosti bijelih lisica, zabilježeno u eksperimentu, govori u prilog djelovanju alelnih mutacija, jer su u ovom slučaju bijele lisice genotipske spojeve dvije dominantne mutacije jednog gena, obje s recesivnim letalnim učinkom. Teško je očekivati smanjenje vitalnosti kada dva različita gena interaguju (komplementarnost). Stoga su zaključili da su bijele, bjelolike, gruzijsko bijele, platinaste i srebrno-crne boje kod lisica kontrolirane nizom alela jednog gena, od kojih su tri dominantna s recesivnim letalnim efektom. Genotipovi lisice: bijela A 1 / A 2, A 1 / A 3, A 2 / A 3; bijelo lice A 1/a; platina A 3/a; gruzijsko bijela A 2/a; srebrno-crni a/a (prema Belyaev et al., 1973).
Treba naglasiti da sa višestrukim alelizmom u slučaju heterozigotnosti originalnih oblika, maksimalni broj fenotipskih klasa u split-u ne može biti 3, već 4, kao u gore opisanom slučaju. U populaciji se broj mogućih genotipova sa višestrukim alelizmom povećava mnogo puta; može se odrediti formulom: 1 / 2n (n+1), gdje je n broj alela. Na primjer, ako postoji 7 alela za lokus, broj mogućih genotipova u populaciji će biti 28: 1 / 2 × 7 × 8 = 28.
Utjecaj recesivnih smrtonosnih mutacija na segregaciju
Recesivni autosomni i spolno vezani letjeli, uzrokujući smrt homozigota u ukrštanju heterozigota za smrtonosne, mogu uticati na cijepanje gena vezanih za smrtonosno.U ovom slučaju, omjer potomaka u split-u ovisi o udaljenosti između gena koji se proučava i smrtonosnog, kao i o tipu heterozigota - u cis- ili transpoziciji geni se uvode u heterozigot. Da bi se identificirali smrtonosni, obično se izvode različita probna ukrštanja. Primjer analize letenja je problem br. III.4.
Problem br. III.4
U liniji Drosophila iz reda br. 100, koja sadrži inverzije, polovina ženki je imala sivu, pola žutu boju tijela, a žute ženke su se pokazale sterilne. Svi mužjaci ove linije bili su žute boje. Omjer ženki i muškaraca bio je drugačiji od normalnog, više kao podjela 2♀♀:1♂♂. Pretpostavlja se da je nedostatak mužjaka u liniji br. 100 vjerovatno posljedica prisustva sivih ženki na X hromozomu, koje su očigledno heterozigotne – linija se dijeli na sive i žute ženke. Da bi se utvrdila genetska struktura ove linije i testirala pretpostavka o prisutnosti letalnog u X hromozomu ženki, izvršeno je ukrštanje čiji su rezultati prikazani u nastavku.
Recipročni krstovi

Sive ženke od F 1 su bili individualno ukršteni sa žutim mužjacima iz linije br. 100.

Analiza
Na osnovu analize zapisati ćemo šeme svih prelaza.

Predložene hipoteze prilično dobro objašnjavaju sve rezultate. Međutim, ostaje nejasno pitanje razloga steriliteta žutih ženki u redu broj 100. Pozivamo čitaoca da razmisli o ovom pitanju i predloži neku hipotezu za njegovo objašnjenje.
U biljke mnoge recesivne mutacije povezane su s nedostatkom ili odsutnošću hlorofila, što dovodi ili do smanjenja vitalnosti biljaka ili do njihove smrti u različitim fazama razvoja. To uzrokuje odstupanja u cijepanju, a također čini potrebnim da se uzme u obzir cijepanje ne samo kod sadnica, već iu kasnijim fazama razvoja kako bi se utvrdio udio uginuća biljaka i priroda nasljeđivanja osobine. Dakle, kukuruz je homozigotan za gen wd (nedostatak bijele boje) imaju bijele sadnice (rascjep na sadnice 3/4 zelene: 1/4 bijele). Međutim, nakon 1-3 sedmice, sve bijele biljke odumiru nakon što se potroše rezerve hranjivih tvari sjemena i u najnaprednijim fazama razvoja biljke, rascjep nestaje - 3 zelene: 0 bijele. Slične mutacije poznate su u grašku, ječmu, raži, pšenici itd.
Ostale mutacije uzrokuju smrt samo dijela jedinki u određenoj fazi razvoja, što dovodi do smanjenja udjela recesiva u rascjepu i promjene odnosa fenotipova - 4:1, 5:1 itd. Ovi omjeri variraju, jer, po pravilu, održivost takvih mutanata u velikoj mjeri zavisi od uslova.
Kod ljudi smanjena vitalnost i smrtnost zbog djelovanja recesivnih mutacija manifestira se u različitim periodima embriogeneze iu različitim fazama razvoja. Razlozi za smanjenje vitalnosti i smrtonosni učinak mogu biti povezani i s mutacijama gena i s kromosomskim abnormalnostima. Citogenetska analiza pobačenih embrija omogućava nam da utvrdimo uzrok smrti mnogih od njih. U prosjeku, zbog hromozomskih aberacija u svim fazama trudnoće, dolazi do više od 42% spontanih pobačaja: značajan dio novorođenčadi sa hromozomskim aberacijama umire tokom prve i narednih godina života.
Među smrtonosnim genskim mutacijama koje dovode do smrti fetusa ili smrti u djetinjstvu mogu se navesti recesivne mutacije koje uzrokuju talasemiju, anemiju srpastih stanica, cističnu fibrozu, kongenitalnu ihtiozu, anencefaliju (odsutnost mozga), fenilketonuriju itd.
Za proučavanje smrtonosnih ili mutacija koje smanjuju vitalnost kod ljudi, široko se koriste citogenetske i biohemijske metode analize, proučavanje strukture i aktivnosti enzima u zdravlju i bolesti, kao i kod heterozigotnih nosilaca; hromatografija, različite vrste elektroforeze.
Smrtonosni geni su mutacijski geni koji uzrokuju smrt pojedinca prije nego što dostigne seksualnu zrelost. Mogu biti dominantni, recesivni ili vezani za spol. Obično pokazuju svoj učinak u homozigotnom stanju, au heterozigotnom stanju smanjuju vitalnost. Penetracija je sposobnost gena da se manifestuje fenotipski, izražena u % i može biti potpuna ili nepotpuna. Potpuna – kod svih pojedinaca u populaciji koji imaju dati gen, manifestuje se kao osobina. Nepotpuni - neki pojedinci imaju gen, ali se ne manifestiraju spolja. Ekspresivnost je stepen ispoljavanja osobine, tj. ista osobina je izražena sa različitim intenzitetom kod različitih pojedinaca.
U prisustvu različitih smrtonosnih gena, organizmi umiru u različitim fazama razvoja. Po pravilu, smrtonosni efekat takvih gena je recesivan, tj. manifestuje se samo kada su u homozigotnom stanju. Kada se pojave mutacije sa dominantnim smrtonosnim efektom, organizam umire bez stvaranja potomstva.
Međutim, bilo je slučajeva kada su smrtonosni geni, koji uzrokuju vidljive promjene u heterozigotnom stanju, čak postali korisni sa ekonomske tačke gledišta. Tako među ovcama karakulske pasmine postoje životinje s prelijepom srebrno-sivom bojom kože, koja se cijeni skuplje od običnog crnog karakula. Prilikom ukrštanja sivih ovaca sa sivim ovnovima pokazalo se da su uvijek heterozigoti. Proučavanjem uzroka ove pojave utvrđeno je da je među sivim jagnjadima dobivenim takvim križanjem otprilike 1/3, odnosno oko 25% ukupnog potomstva, oboljelo od kroničnog timpanitisa i umrlo. Uzrok bolesti su poremećaji u aktivnosti parasimpatičkog nervnog sistema. Ukrštanjem sivih ovnova sa crnom ovcom ili sive ovce sa crnim ovnovima, potomci su dali 50% sivih i 50% crnih jagnjadi, a siva jagnjad nisu oboljela. Pokazalo se da je u homozigotnom stanju gen koji je izazvao razvoj sive boje imao recesivni letalni učinak.
Lisice imaju vrijednu boju krzna - platinastu, uzrokovanu dominantnim genom koji u homozigotnom stanju uzrokuje smrt embriona u ranoj fazi razvoja. U takvim slučajevima se menja odnos fenotipova u drugoj generaciji, jer od tri dominantna oblika jedan (homozigot za smrtonosni gen) umire, usled čega odnos fenotipova postaje 2:1.
Slično cijepanje zabilježeno je i kod zrcalnog šarana, kod kojeg je uočeno nasljeđivanje nerazvijenosti ljuski: očuvan je samo na srednjoj liniji tijela, ostatak je bio gol, zbog čega se naziva linearnim. Kada se linearni šarani međusobno ukrštaju, potomci se uvijek dijele u omjeru 2 linearna: 1 sa normalnim ljuskama, budući da embrioni homozigotni za gen koji uzrokuje nerazvijenost ljuski umiru u ranim fazama razvoja.
Kod pilića homozigotnih za alel koji uzrokuje uvijanje perja, nepotpuni razvoj perja ima nekoliko fenotipskih efekata. Ovi pilići imaju nedovoljnu toplinsku izolaciju i pate od hlađenja. Kako bi nadoknadili gubitak topline, razvijaju brojne strukturne i fiziološke adaptacije, ali su te adaptacije neučinkovite i smrtnost je visoka među takvim kokošima.
Učinak smrtonosnog gena jasno se vidi u nasljeđivanju boje dlake kod miševa. Divlji miševi obično imaju sivo krzno, poput agoutija; ali neki miševi imaju žuto krzno. Ukrštanja žutih miševa proizvode i žute miševe i agouti u omjeru 2:1. Jedino moguće objašnjenje za ove rezultate je da je žuta boja dlake dominantna kod agoutija i da su svi žuti miševi heterozigoti. Atipični mendelovski odnos objašnjava se smrću homozigotnih žutih miševa prije rođenja. Nekropsije trudnih žutih miševa ukrštenih sa žutim miševima otkrile su mrtve žute mladunce u njihovim maternicama. Kada bi se ukrštali žuti miševi i agouti, tada ne bi bilo mrtvih žutih miševa u maternicama trudnih ženki, jer takvim ukrštanjem ne može biti potomaka homozigotnih za gen žute vune.
Promjena fenotipske segregacije u omjeru 3:1 u drugoj generaciji monohibridnog ukrštanja povezana je sa različitom održivošću F2 zigota. Različita održivost zigota može biti posljedica prisustva smrtonosnih gena. Smrtonosni je gen koji uzrokuje poremećaje u razvoju organizma, što dovodi do njegove smrti ili deformiteta.
Proučavanje kongenitalnih anomalija pokazalo je da je s različitim smrtonosnim genima smrt jedinki različita i može se dogoditi u različitim fazama razvoja.
Prema klasifikaciji koju je predložio Rosenbauer (1969), geni koji uzrokuju smrt 100% jedinki prije nego što dostignu zrelost nazivaju se smrtonosnim, više od 50% subletalnim (poluletalnim), a manje od 50% subvitalnim. Međutim, treba napomenuti da je ova podjela u određenoj mjeri proizvoljna i ponekad nema jasne granice. Primjer je spolno vezana golotinja kod pilića. Gotovo polovina golih pilića ugine u posljednja 2-3 dana inkubacije. Od izleženih pilića, otprilike polovina ugine prije 6 sedmica starosti ako se uzgajaju na temperaturi od 32-35 °C. Ali ako se temperatura u leglu poveća za 5,5 °C, tada će uginuti znatno manje golih pilića. 4-- Nakon 5 mjeseci, goli pilići rastu rijetko perje i već mogu izdržati prilično niske temperature.
U prirodnim uslovima, ova mutacija će verovatno biti smrtonosna i dovesti do 100% smrtnosti ptica. Gornji primjer pokazuje da priroda manifestacije polusmrtonosnog gena može u velikoj mjeri ovisiti o uvjetima okoline.
Smrtonosni geni mogu biti dominantni ili recesivni. Među prvim smrtonosnim faktorima otkriven je alel koji je uzrokovao žutu boju miševa. Gen za žutu boju je dominantan (Y). Njegov recesivni alel (y) u homozigotnom stanju uzrokuje pojavu crne boje. Međusobno ukrštanje žutih miševa dalo je dva dijela žutih miševa i jedan dio crnih, odnosno rezultat je bio 2:1, a ne 3:1, kako slijedi iz Mendelovog pravila. Pokazalo se da su svi odrasli miševi heterozigoti (Yy). Prilikom međusobnog ukrštanja trebali su proizvesti jedan dio homozigotnog potomstva za žutu boju (IT), ali ono umire u embrionalnom periodu, dva dijela heterozigota (Yy) će biti žuta, a jedan dio homozigota za recesivna osobina (yy) će biti crna. Shema križanja izgleda ovako:
Na isti način, siva boja dlake nasljeđuje se kod karakulskih ovaca (Sokolskie, Malich, itd.), platinasta boja kod lisica, distribucija ljuske kod linearnih šarana itd.
Smrtonosni geni su u većini slučajeva recesivni i stoga mogu dugo ostati latentni.
Životinja koja je potpuno zdrava i normalna po fenotipu može biti nosilac smrtonosnog gena, čiji se učinak otkriva tek pri prelasku u homozigotno stanje. Smrtonosni geni najčešće prelaze u homozigotno stanje tokom inbridinga. U praksi stočarstva pri uzgoju konja zabilježen je slučaj uginuća 25 ždrebadi 2.-4. dana nakon rođenja od deformiteta rektuma - odsustva anusa (Atresia ani). Ispostavilo se da su svi pastuvi i kobile koje su rodile tako nenormalna ždrebad potekle od istog pastuha. Bio je heterozigotan za smrtonosni gen (LI). U početku je ovaj pastuh, ukrštanjem sa normalnim kobilama (LL), dao potomstvo normalnog fenotipa, ali u smislu genotipa polovina potomaka je bila normalna (LL), a polovina heterozigota (LI), koja je nosila recesivna sklonost (0 smrtonosni gen. U inbridingu heterozigotnih životinja (Y x Y) pojavila su se neka ždrebad, homozigotna po smrtonosnom genu (II), sa deformitetom rektuma. Svi su umrli.
Poznato je na desetine anomalija kod domaćih životinja, čija je pojava povezana s recesivnim ili dominantnim genskim mutacijama. Ove anomalije se javljaju u pojedinim populacijama sa različitom učestalošću, što zavisi od brzine procesa mutacije, sistema uzgoja životinja itd. Poznavanje specifičnih oblika kongenitalnih anomalija kod životinja svake vrste, kao i učestalosti njihovog ispoljavanja u pojedinih pasmina, neophodna je veterinarskim specijalistima za selektivnu prevenciju širenja genetske patologije.
Anomalije kod goveda. Biološke karakteristike ove vrste životinja su kratkodlake i relativno kasno sazrevaju. Krava obično rodi jedno tele, koje dostiže polnu i fiziološku zrelost tek za 1,5 godinu, tako da je period između prvog teljenja majke i ćerke u prosjeku 5 godina. Kao rezultat toga, pojava abnormalnog potomstva u stadu može značajno smanjiti razinu reprodukcije i intenzitet uzgojne selekcije stoke. U goveda je proučavan širok spektar kongenitalnih šamalija, određenih smrtonosnim, poluletalnim i subvitalnim genima. 46 anomalija uključeno je u Međunarodnu listu smrtonosnih mana pod šifrom A (tabela 43). Relativna učestalost određenih vrsta abnormalnosti u svakoj rasi ili populaciji može varirati. Kod kostromske pasmine, prema našim podacima, najčešće zabilježena genetska anomalija glave je skraćivanje čeljusti (tabela 44), kod jaroslavske pasmine - sindaktilija, kod holmogorske - mišićne kontrakture, kod crno- Bijela - pupčana kila. Abnormalnosti centralnog nervnog sistema bile su najčešće (21%) kod goveda u Njemačkoj.
Drugo mjesto po učestalosti registracije (14%) zauzela je složena anomalija - kombinacija pupčane kile s rascjepom abdomena i fetusa u cjelini. Učestalost anomalija, odnosno procenat abnormalnog potomstva u ukupnom broju, unutar pojedinih populacija takođe može biti veoma različita i, prema prosečnim procenama, ne prelazi 1%. Međutim, ovaj pokazatelj ovisi o potpunosti i tačnosti evidentiranja anomalija. Tako su u Njemačkoj, nakon organiziranja čistog računovodstva, zaključili da se učestalost anomalija povećala nekoliko puta. Pitanje je: da li su sve anomalije podložne vizuelnom posmatranju? Očigledno ne sve. Tako je kod kostromske rase prosečna učestalost svih oblika anomalija u periodu od 12 godina bila 1,15%. Učestalost ukupnog mortaliteta legla (pobačena, mrtvorođena, abnormalna, mrtva telad bez vidljivih nedostataka) na ovoj farmi iznosila je 10,2%. Određeni dio ovog mortaliteta povezan je i s mutacijama gena koje uzrokuju ne morfološke defekte, već metaboličke poremećaje i druge anomalije, čija je identifikacija moguća samo posebnim metodama.
Proizvođači mogu igrati posebnu ulogu u širenju genetskih abnormalnosti kako kod goveda tako i kod drugih životinjskih vrsta. Od svakog oca, veštačkom oplodnjom, godišnje se mogu dobiti stotine i hiljade potomaka. Tako je od jednog bika u inostranstvu dobijeno 100 hiljada teladi. Ako se pokaže da je takav otac nosilac genske mutacije, brzo će se proširiti rasom. Evo nekoliko primjera iz brojnih činjenica opisanih u literaturi. Kao rezultat intenzivne upotrebe bika princa Adolfa, dovezenog u Švedsku, i naknadnog spontanog parenja s njim, učestalost bezdlake kod nekih švedskih krda bila je preko 5%. Ista situacija nastala je u Švedskoj nakon uvoza bika Gallus, za koji se ispostavilo da je heterozigotni nosilac gena koji uzrokuje odsustvo udova.
Kod potomaka pojedinačnih bikova crno-bijelog i šarolea u SAD-u i Njemačkoj zabilježeni su slučajevi rođenja patuljastih teladi sa učestalošću od 23,3 i 22,2%, respektivno. U prvom U Čehoslovačkoj, prilikom ispitivanja potomaka 166 potomaka, utvrđeno je da su 43 nosioci smrtonosnih gena. Kod jednog bika - nosioca dominantne anomalije "rascjep usne" - mana se manifestirala kod 44% bikova i 71% junica iz njegovog potomstva.
Kod kostromske pasmine analizirali smo širenje skraćivanja donje vilice i izgleda poput mopsa preko bika Burkhan, koji je i sam imao defektnu telad u svom potomstvu; njegovi sinovi, unuci, praunuci, ženski potomci su takođe dali abnormalno potomstvo (Sl. 57). Većina teladi je dobijena od inbreedinga i parenja roditelja sa normalnim fenotipom i zajedničkim pretkom. Stoga možemo zaključiti da ova anomalija ima recesivni način nasljeđivanja. Na slici je vidljivo da je najveći broj defektnih teladi registrovan kod potomaka bika Zheton 3501 (unuka bika Burkhan) pri korištenju na komercijalnoj farmi, gdje su neke od krava imale isti recesivni gen u svom genotipu.
Anomalije kod svinja. Međunarodna lista smrtonosnih mana kod svinja uključuje 18 genetskih abnormalnosti. Najveći dio njih uzrokovan je autosomno recesivnim genima (tabela 45). Genetske abnormalnosti mogu igrati značajnu ulogu u patologiji svinja. Pogledajmo nekoliko takvih primjera. U Španiji, istraživanje 23.449 prasadi iz 2.399 legla dobijenih od nerastova rasa Duroc, Yorkshire, Hampshire i White Chester, pokazalo je, respektivno, 6,21; 6.02; 9.66; 2^62% abnormalnih legla.
Prema Olivijeu (1979), 7 genetskih abnormalnosti kože, 17 skeleta, 3 očiju, 13 neuromišićnih, 6 krvi, 6 hormonskog metabolizma, 5 probavnog sistema i 9 abnormalnosti genitourinarni sistem opisani su kod svinja. Glavne anomalije bile su kriptorhizam, hernije, pseudohermafroditizam itd. Autor studije smatra da su ove anomalije rezultat djelovanja jednog gena u različitim fazama formiranja embrija.
U Danskoj je tokom dvogodišnjeg perioda proučavano 6.669 uginulih prasadi iz 2.936 legla kako bi se utvrdila priroda i učestalost urođenih anomalija. Različite anomalije otkrivene su kod 1,4% rođenih prasadi, odnosno 6,2% ubijenih prije odbića. Prilikom obdukcije kod 25,9% abnormalnih prasadi utvrđeni su nedovoljno razvijeni zalisci, nezatvaranje anusa, subaortna stenoza, ektopija srca i druge mane kardiovaskularnog sistema. Kod 23,4% prasadi utvrđeni su različiti razvojni poremećaji motoričkog sistema. Anomalije centralnog nervnog sistema otkrivene su kod 5,9% prasadi, uključujući bifurkaciju mozga i cerebralnu hidrokelu. Prekomjerni rast rektuma, tankog crijeva ili njihov nepotpuni razvoj utvrđen je kod 30% prasadi, a razne kile i ascitesi kod 6,8%. Rascjep usne, rascjep nepca, rinocefalitis i druge anomalije facijalnog dijela glave nađeni su kod 6,1%; hermafroditizam, rascjep uretera, hidrokela bubrega i uretre - kod 1,7% prasadi. Ove anomalije su se pojavile u potomstvu pojedinačnih bića tokom inbreedinga, što ukazuje na nasljednu prirodu njihove pojave.
Fridin i Newman dobili su vrlo uvjerljive dokaze o nasljednoj prirodi kriptorhizma kod svinja. Prema njihovim podacima, u Kanadi se jednostrani i bilateralni kriptorhizam svake godine uočava kod 1-2% svih nerastova koji ulaze na tržište. Autori su ukrštali kriptorhide sa njihovim majkama i punim sestrama. Potomci od takvih ukrštanja su međusobno pareni. Kao rezultat takve selekcije i selekcije, učestalost kriptorhizma kod pokusnih životinja jorkširske pasmine porasla je u prosjeku na 42,9%, a posebno kada su korištena dva proizvođača. Prilikom pregleda prasadi u Sjedinjenim Državama u jednoj godini pronađeno je oko 400 tisuća životinja sa skrotalnom hernijom.
Opažanja pokazuju da je uzrok smanjene plodnosti kod nerastova često hipoplazija testisa. Učestalost ove anomalije, prema istraživačima iz Njemačke, iznosila je 19,6%. 30 takvih nerastova ostavljeno je za reprodukciju
Va, svaka od njih pokrivala je od 4 do 40 matica (ukupno 439 grla), ali samo četiri su rodile potomstvo. Analiza je pokazala da su patološki oblici sperme kod ovih nerastova 80-100%. Svih 30 abnormalnih životinja imalo je zajedničke pretke, što ukazuje na nasljednu prirodu hipoplazije testisa i defekta spermiogeneze.
Prisustvo bradavica kratera kod svinja je jedan od ozbiljnih nedostataka, jer prasad od njih ne dobija mlijeko. Prema Bavarskom institutu za stočarstvo (Njemačka), učestalost ove anomalije u njemačkim landrasima bila je 6,6%. Kao što su primijetili P. N. Kudryavtsev i saradnici (MVA), broj svinja s neaktivnim sisama kratera se posljednjih godina povećao. Broj takvih bradavica kreće se od 1 do 8. Prasadi koja dobiju bradavice u obliku kratera uginu.
Krateritet je osobina uzrokovana jednim autosomno recesivnim genom. Ovo su eksperimentalno testirali P. N. Kudryavtsev et al. Nakon što su prethodno identificirali nerastove i nazimice koji su nosioci anomalija (KchKch), normalne ali heterozigotne jedinke (KchKch), koje proizvode prasad sa kraterom u potomstvu, i normalne homozigotne svinje (KhKch), autori su izvršili ukrštanje između ovih grupa životinja. . U prvoj varijanti ukrštano je 27 normalnih homozigotnih brana sa 15 nerastova. Svih 258 potomaka bilo je normalno. U drugoj varijanti, gdje je jedan od roditelja bio homozigot (KchKch), a drugi heterozigot (Kchkch), svi prasadi su također bili normalni. U trećoj varijanti ukršteno je 13 heterozigotnih nerastova sa 16 heterozigotnih mama. Od 168 rođenih prasadi, njih 39 (23,2%) imalo je bradavice kratera. I konačno, u četvrtoj opciji, jedan od roditelja je bio homozigot, a drugi heterozigot. Dobili su 170 prasadi, od kojih je 86 (50,5%) imalo normalne bradavice, a 84 (49,5%) bradavice sa kraterima. Rezultati ovog eksperimenta dokazuju recesivni način nasljeđivanja kratera bradavice kod svinja.
Abnormalnosti kod ovaca. Opisano je oko 90 urođenih anomalija kod ovaca. Prema Dennisu i Leipoldu, većina poznatih genetskih defekata kod ovaca uzrokovana je monogenim autosomno recesivnim načinom nasljeđivanja (tabela 46). Najčešći kraniofacijalni defekti kod ove vrste životinja su kraniofacijalni defekti, posebno agnatija, kao i zakrivljenost prednjih udova, mikroagnatija, hermafroditizam, kriptorhidizam, hipospadijaza, prognatija, analna atrezija, mikrotija, entropija, potorogliza, art. Analiza je pokazala da se 55,4% defekata odnosi na mišićno-koštani sistem, 12,7 - na probavni sistem, 9,7 - na kardiovaskularni sistem, 7,1 - na urogenitalni sistem, 6 - na centralni nervni sistem, 3.5 - na anomaliju od ligamenata, 3,2 - na abdominalni, 1,5% - na endokrini sistem. Iako je učestalost pojedinačnih nedostataka niska, kumulativni doprinos svih anomalija može uzrokovati štetu na farmama. U Novom Zelandu, zemlji s razvijenim ovčarstvom, incidencija smrtonosnih mana iznosila je oko 1% uginulih jagnjadi. U SAD, smrtonosni defekti u prve 2 sedmice života uočeni su kod 11,4% jagnjadi.
Prosječna stopa embrionalnog mortaliteta kod ovaca je 20%. Ovo ukazuje da mnogi neidentifikovani smrtonosni geni mogu biti aktivni tokom ovog perioda.
Pojedinačne genetske abnormalnosti kod ovaca mogu biti široko rasprostranjene. Tako je u Bugarskoj, u krdima merino ovaca, uočen visok mortalitet jagnjadi u ranom postnatalnom periodu. Nastala je kao rezultat činjenice da jagnjad nisu dobijala mlijeko zbog nedostataka vimena njihovih majki: od hipoplazije sa malim ostacima žljezdanog tkiva do njegovog potpunog odsustva. Učestalost ove nasljedne anomalije u različitim stadima kretala se od 6 do 40%.
Kod ovaca se često opaža kriptorhizam, koji je kombiniran s takvom sekundarnom seksualnom karakteristikom kao što je polnost. Ovnovi ovnovi imaju nisku plodnost. Strogom selekcijom uzgoja bilo je moguće stvoriti tip ovnova s normalnom plodnošću, međutim, među njima se nalaze i abnormalne jedinke.
Anomalije kod ptica. Ptice, prvenstveno kokoške, najviše su proučavane u odnosu na genetiku abnormalnosti. Međunarodna lista smrtonosnih mana uključuje 45 anomalija kod pilića, 6 kod ćurki i 3 kod pataka. Najčešće abnormalnosti kljuna su (kljun papagaja, ukršteni kljun). Njihova učestalost, prema William et al., je 1,1% gubitaka tokom inkubacije jaja od pilića White Leghorn i Rhode Island. Abnormalnosti kljuna su takođe česte kod pataka.
Max Gibbon i Shakelfurd opisali su anomaliju pri ukrštanju bijelih leghorna s buttercamps i bantamima i naknadnim razmnožavanjem "u sebi" - polidaktilijom. Osim toga, kod pilića je uočena sindaktilija i pernate noge. Učestalost sindroma bila je 16,8%. Prilikom ukrštanja abnormalnog F2 pijetla sa fenotipski normalnim kokošima, uočen je podjel - pola normalne i pola abnormalne jedinke (1:1). Utvrđeno je da ovaj kompleks osobina kontroliše jedan autosomni gen, koji ima polu-smrtonosno dejstvo, budući da je embrionalno i postembrionalno preživljavanje abnormalnih pilića veoma nisko.
Anomalije kod konja. Od nasljednih anomalija kod konja, 10 je uključeno na Međunarodnu listu smrtonosnih mana. Među njima su 3 anomalije skeleta, 2 reproduktivnog sistema, 2 bubrega i mišića, po jedna anomalija crijeva, nervnog sistema i organa vida.
Kod konja vučnih rasa češća je atrezija debelog crijeva. Prevalencija ove anomalije zabilježena je kod potomaka pastuha Superba pasmine Percheron. Također je opisana kod ždrebadi čistokrvne jahaće rase istočnoafričkog porijekla. Nesavršena epiteliogeneza zabilježena je kod vučnih pasmina. Ataksija, nazvana Oldenburg, pronađena je kod ždrebadi Oldenburg u Njemačkoj. Širi se u liniji 9. Jedna od najčešće zabilježenih anomalija kod konja je pupčana kila. Nalazi se kod lakih i teških pasmina.
Kod konja je poznato nekoliko drugih genetskih i nasljedno-ekoloških anomalija. Tako u SAD-u konji nekoliko pasmina imaju ždrebad sa osebujnom bijelom mrljom koja se naziva “overo”. Prilikom ukrštanja konja tipa "overo" rađaju se ždrebad s ružičastom kožom, u kojoj se uočava hipoplazija crijevnog trakta i izoeritroliza, kao i kolike koje dovode do smrti.
U Engleskoj su registrovane i proučavane životinje s poremećenom koordinacijom pokreta - "bolest kolebanja" na konjima polukrvnih jahačkih pasmina. Utvrđena je genetska predispozicija za ovu anomaliju.
Nasljednost dermatoza ekstremiteta kod konja je dokazana. Defekti s nasljednom predispozicijom su često javljanje kronične deformirajuće upale skočnog zgloba – špara, tzv. “štaka” kod ždrebadi, kao i kronična aseptična upala koronalnog bloka kopita, koja se uočava uglavnom kod trkaćih konja i trkaćih konja. konji.









- Pokretna imovina stečena nakon 01
- Porez na imovinu stečenu nakon 01
- Morski konjic - izvještaj s porukom Koliko dugo morski konjic postoji na zemlji
- Keksi od vanile sa mliječnom čokoladom
- Recept za palačinke sa kiselim kvascem sa fotografijom
- Istorija pronalaska votke
- Naslijeđe svetih otaca: pravoslavne parabole o životu i moralu i biblijski citati
- Kraljevsko meso, novogodišnji recept za kuvanje kraljevskog mesa Kraljevsko meso sa krompirom u rerni
- Bijeli kvas za okrošku Napravite okrošku kvas kod kuće
- Hemija (struktura atoma) (prezentacija)
- Tradicija i običaji Engleske koji će vas iznenaditi
- Nasljednost i razvoj Da li je naslijeđe toliko važno?
- Harry Potter knjige na engleskom
- Organizacioni faktori koji uzrokuju stres
- Recept za dinstani kupus u sporom šporetu: jednostavan, brz i ukusan
- Osmica štapova, opis i karakteristike karata
- Medaljoni od bifteka i lososa
- Sažetak individualnog logopedskog časa o automatizaciji zvuka Rezime automatizacije zvuka l u rečenicama
- Nižnji Novgorodski državni pedagoški univerzitet nazvan po
- Svi oni koji su razapeli Hrista umrli su strašnom smrću.Da li je Pontije Pilat zaista bio tamo?