Recesivni, dominantni, letalni geni, mutacije. III.2.8
Do sada smo ljudsku genetiku razmatrali prema tipovima nasljeđivanja, a sada ćemo se osvrnuti na jednu vrlo važnu kategoriju gena, naime letalne i subletalne gene. Letalni i subletalni geni su klasa mutacija s izrazito izraženim negativnim učincima na razvoj. Kao što ćemo kasnije vidjeti, mutacije u mnogim slučajevima pogoršavaju i uništavaju nasljeđe osobe. Ovaj


pogoršanje mutacijama posebno je izraženo pod utjecajem letalnih gena.
Letalni i subletalni aleli mogu biti dominantni ili recesivni. U slučaju dominantnog letaliteta dijete umire u prenatalnom razdoblju ili u ranom djetinjstvu. Posve je jasno da se takvi geni ne mogu naslijediti. Njihova prisutnost ukazuje na pojavu mutacija. Ukupna učestalost takvih dominantnih letala nije tako mala. Njihov je broj posebno velik pod utjecajem zračenja. U genetici miševa i Drosophile, dominantni letali naširoko se koriste pri analizi ovisnosti učestalosti mutacije o dozi i pri analizi drugih značajki mutagenih učinaka. U prisutnosti subletalnih, inače nazvanih polu-letalnih gena, dijete umire u ranom djetinjstvu ili nešto kasnije, ali u pravilu prije puberteta. Primjer dominantne subletalne mutacije je epiloja, bolest koju karakterizira djelovanje čitavog sindroma negativnih simptoma. Djeca s epiloyijom imaju patološke izrasline na koži, karakteriziraju ih mentalna retardacija, epilepsija, tumori srca, bubrega i drugih organa. U rijetkim slučajevima pacijenti s epilojom prežive i vjenčaju se sa zdravim supružnikom. Polovica djece iz takvog braka ispada bolesna od kongenitalne epiloije (slika 131).
Još jedan upečatljiv primjer subletalne dominantne mutacije je bolest retinoblastom. Ovaj


dominantni gen uzrokuje kancerogeni tumor u oku, što gotovo uvijek dovodi do smrti djeteta u ranoj dobi.
Ipak, najveći dio kongenitalnih deformacija otpada na recesivne letale i subletale. U ovom slučaju, recesivni štetni geni skriveni su u genotipu naizgled zdravih roditelja, međutim, kada se dva takva heterozigota vjenčaju, oko 25% njihove djece ispada bolesno.
Slika 132 prikazuje glavne vrste križanja u ljudskim populacijama koja se provode pri nasljeđivanju recesivnih letalnih i subletalnih gena. Na kvadrat 1 prikazuje tip brakova između genetski normalnih ljudi. Kod oba roditelja i svih njihovih potomaka, svi aleli su predstavljeni normalnim genima (alel +). Na kvadrat 2 prikazuje brak između genetski zdrave osobe i heterozigota (simbol a). U ovom slučaju, negativni alel se prenosi na polovicu potomaka, ali ostaje skriven. Na kvadrat 3 genetski zdrava osoba udaje se za bolesnu osobu (simbol aa), sva djeca su naizgled zdrava, ali u heterozigotnom stanju nose subletalnu mutaciju A. Na kvadrat 4 heterozigot se ženi pacijentom; polovica njihove djece je naizgled zdrava, ali nosi subletalnu mutaciju, druga polovica je bolesna. Na kraju, kvadrat 5 u slučaju braka dvoje bolesnih (simboli aa) svi će potomci biti bolesni. Kongenitalne bolesti uzrokovane djelovanjem recesivnih letalnih i subletalnih gena uključuju dječju amaurotičnu idiotiju, u kojoj dolazi do degeneracije moždanog tkiva, sljepoće i smrti djeteta; kongenitalna patologija kože s prisutnošću
duboke krvareće pukotine - ihtioza (Sl. 133); kongenitalna dječja paraliza i niz drugih kongenitalnih bolesti.
Mnogi rani spontani pobačaji uzrokovani su curenjem smrtonosnih genotipova.
Poznate su dominantne mutacije koje kod heterozigota određuju neko više ili manje neutralno svojstvo, koje, međutim, imaju letalni učinak u homozigotnom stanju. Na primjer, pigmentacijski alel kseroderme u heterozigotnom stanju uzrokuje ozbiljne pjegavosti. Međutim, homozigoti razvijaju xeroderma pigmentosum. Ovo je subletalna bolest (slika 134), kod koje djelovanjem svjetla dolazi do oštećenja kože, što zatim dovodi do malignog rasta.
Jedan od alela kod osobe u heterozigotnom stanju uzrokuje višak kolesterola u krvi. Međutim, djeca homozigotna za ovaj alel razvijaju ozbiljne subletalne abnormalnosti.
U svim tim slučajevima imamo klasičnu sliku nasljeđivanja dominantnih s recesivnim letalnim učinkom, prethodno utvrđenu u pokusima s drozofilom, miševima i drugim organizmima. Na primjer, miš ima dobro poznati alel A Y, što uzrokuje žutu boju kože kod heterozigota. Međutim, svi pokušaji da se dobije čista rasa žutih miševa ostali su neuspješni. Prilikom križanja dva žuta miša, u prosjeku, dva žuta miša uvijek daju jednog različite boje, na primjer crnog. Priroda ovog fenomena objašnjena je činjenicom da se gen za dominantnu žutu boju pokazao
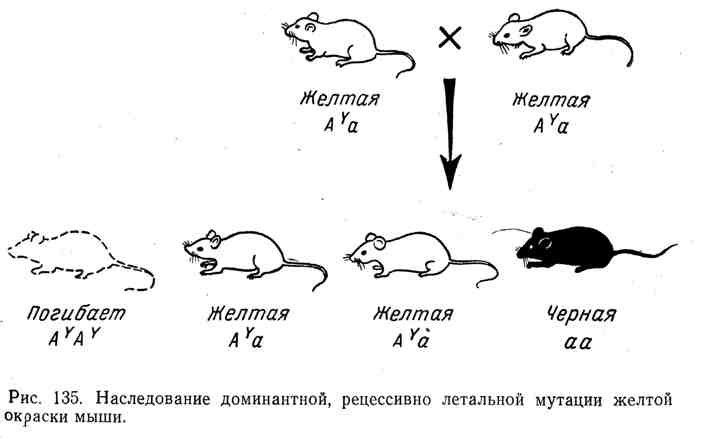
recesivno smrtonosno. Svi homozigoti( A D A D) umro. Žuti miš je uvijek heterozigot, zajedno s alelomA Y sadrži drugi alel, npr. A(Crna boja). Tijek nasljeđivanja pri križanju heterozigota A Y a i cijepanje prema formuli 2:1 prikazani su na slici 135.
- Izvor-
Dubinin, N.P. Horizonti genetike / N.P. Dubinjin. – M.: Obrazovanje, 1970.- 560 str.
Broj pregleda: 121
Nejednaka sposobnost preživljavanja zigota različitih genotipova može biti povezana s dominantnim ili recesivnim letalnim mutacijama koje nastaju u gametama roditeljskih jedinki. Mogu se ostvariti u različitim fazama embriogeneze ili u postembrionalnom razdoblju. Naravno, smrt nekih zigota utječe na prirodu cijepanja.
Utjecaj na cijepanje dominantnih gena s recesivnim letalnim učincima
Dominantni geni s recesivnim letalnim učincima imaju pleiotropni učinak; s jedne strane, oni određuju dominantnu prirodu manifestacije bilo kojeg svojstva u heterozigotu, s druge strane, uzrokuju smrt jedinki u homozigotu, tj. manifestiraju svoj smrtonosni učinak kao recesivni. Takvi su geni poznati kod mnogih životinja - žuta boja kod miševa, kratke noge kod kokoši, linearna ljuskavost kod šarana, platinasta boja kod lisica, shirazi boja kod ovaca i mnogi drugi. Oni uzrokuju odstupanje od rascjepa 3:1, pretvarajući ga u rascjep 2:1. Analiza segregacija koje uključuju dominantne gene s recesivnim letalnim učincima komplicirana je činjenicom da u relativno malim uzorcima nije uvijek moguće razlikovati podjele 3:1 i 2:1, a statistička metoda testiranja pomoću χ 2 kriterija ne dopušta izbor iz hipoteza. Na primjer, ako se kod međusobnog križanja žutih miševa dobije podjela od 65 žutih: 32 crna, metoda %2 ne odbacuje dvije hipoteze: 1 - podjela u eksperimentu odgovara podjeli od 3:1, χ2 = 3,29, p>0,05; 2 - cijepanje u eksperimentu odgovara cijepanju 2:1, χ 2 = 0,17, p>0,05.
Samo u velikim uzorcima može se razlikovati 2:1 podjela od 3:1.. Kada se sumiraju podaci o nasljeđivanju žute boje kod miševa koje su dobili različiti istraživači, podjela pri međusobnom križanju žutih miševa bila je 2386 žutih: 1235 crnih - ∑ = 3621, χ 2 = 0,96 pri H 0 - 2: 1 ( kod H 0 - 3:1 u ovom slučaju χ 2 = 160!).
Očito je da različite metode genetskog testiranja, primjerice provođenje analitičkih i povratnih križanja, mogu pridonijeti ispravnom rješenju pitanja nasljeđivanja. U gore razmotrenom primjeru to su križevi: žuti od F 1 × crni; žuto od F 1 × žuto P. U prvom slučaju treba postojati cijepanje 1: 1, u drugom - opet 2: 1.
Teže je otkriti da postoji odstupanje i to zbog različite vitalnosti zigota u slučaju kada do smrti nekih zigota dođe u postembrionalnom stadiju. S tim u vezi potrebno je pratiti plodnost i mortalitet podmlatka ukoliko postoji pretpostavka letaliteta. Osnova za pretpostavku letaliteta je nemogućnost dobivanja ujednačenog potomstva pri uzgoju jedinki određenog fenotipa. Primjeri analize dati su u zadacima br. III.2 i III.3.
Problem br. III.2
U potomstvu križanja srebrno-sable minks jedan s drugim u F 1, uvijek se opaža razdvajanje: pojavljuju se srebrno-sable i smeđi pojedinci.
U jednom pokusu, nekoliko legla križanaca srebrnog samura dalo je 93 šteneta srebrnog samura i 43 smeđa, s prosječnom veličinom legla od 3,66 štenaca. Križanjem srebrnog samura sa smeđim nercima dobiveno je 39 smeđih i 41 srebrni samur. U međusobnom križanju smeđih nerca dobiveni su samo smeđi I potomci. Veličina legla u zadnja dva križanja bila je 4,9-5,2 šteneta.
Objasnite cijepanje, odredite genotipove srebrnog samura i smeđeg nerca.
Analiza
Budući da srebrno-sable minkovi uvijek stvaraju segregaciju kada se križaju međusobno i sa smeđim, oni su očito heterozigoti. Da biste odredili broj gena, razmotrite podjele u eksperimentu:
H 0 - razlike u jednom genu, cijepanje 3:1, χ 2 = 3,2, p>0,05. Odstupanje je slučajno, hipoteza se ne odbacuje.

Omjer dobro odgovara podjeli 1:1 za jedan gen, χ 2 = 0,05, p>0,80. Odstupanje je slučajno, hipoteza se ne odbacuje.
Usporedba podataka o plodnosti minkova u različitim križanjima ukazuje na djelomičnu smrt zigota pri križanju srebrno-sable minkova. Možemo pretpostaviti smrt dominantnih homozigota. Tada je genotip srebrno-sable minkova Aa, smeđih - aa, a podjela u križanju nije 3:1, već 2:1 (χ 2 = 0,17, p>0,05). Za konačnu potvrdu ove hipoteze potrebno je izvršiti dodatno križanje jedinki srebrnog samura kako bi se povećao uzorak i statistički testirala hipoteza 2:1. U nekim slučajevima potrebno je provesti složeniju analizu.
Problem br. III.3
Kod križanja platinaste, bijele i gruzijske bijele lisice sa srebrno-crnim, utvrđeno je da platinaste, bijele i gruzijske bijele boje, koje uzrokuju opće slabljenje pigmentacije i pojavu raznih pjegavih lisica, nisu spolne. -povezano. Križanje svakog od ovih mutanata sa srebrno-crnim rezultiralo je segregacijom 2:1, pri čemu je srebrno-crna boja bila recesivna. Posljedično, svaki od njih je kontroliran jednim dominantnim genom s recesivnim letalnim učinkom. O smrti nekih mutanata svjedoče podaci o veličini legla: srebrno-crni imaju 4,5 štenaca po leglu, bijeli 3,5, platinasti i gruzijski bijeli - manje od 3,5.
Kako bi se ustanovilo jesu li te mutacije zahvatile jedan gen ili različite, obavljena su križanja čiji su rezultati dolje navedeni. Test alelizma nije primjenjiv u ovom slučaju, jer su mutacije dominantne:

Objasniti cijepanje, odrediti genotip svih oblika.
Analiza
1. Podjele u križanju 1 i 2 odgovaraju omjeru 1:1:1:1 (χ 2 = 5,83, p>0,10 za križanje 1, χ 2 = 0,55, p>0,90 za križanje 2). One mogu biti rezultat više razloga.
1. Budući da kod križanja 1 i 2 postoje 4 klase cijepanja i omjer 1:1:1:1, može se pretpostaviti da se proučavani oblici razlikuju u dva neovisno naslijeđena dominantna gena s recesivnim letalnim učinkom koji međusobno djeluju prema vrsta komplementarnosti. U ovom slučaju, jedan od gena predstavljen je s dva različita dominantna alela.

2. Segregacija 1:1:1:1 može biti posljedica bliske povezanosti ova dva gena u nedostatku križanja među njima:

(U Punnettovim rešetkama dani su fenotipski radikali - geni koji očituju svoj učinak u fenotipu.)
3. Razlike u boji uzrokuju tri neovisno naslijeđena dominantna gena s recesivnim letalnim učinkom. U prvom (a) i drugom (b) križanju dolazi do cijepanja u dva različita gena.

4. Segregacija 1:1:1:1 može biti rezultat bliske povezanosti triju gena u interakciji u nedostatku crossing overa.

* (Redoslijed gena može biti različit, zadan je proizvoljno.)
5. Segregacija u omjeru 1:1:1:1 može biti rezultat razlika u jednom genu, predstavljenom nizom od četiri alela, od kojih su tri dominantna s recesivnim letalnim učinkom, a četvrti je recesivan:

Kako bi se napravio izbor između ovih hipoteza, napravljen je križ - bijeli sa srebrno-crnim.
Kad bi osobinu kontrolirala dva ili tri gena, tada bi se uz njihovo neovisno nasljeđivanje očekivala pojava četiri fenotipske klase:

Sličan bi rezultat trebao biti i kod križanja bijelih lisica dobivenih od platinastih ili gruzijskih bijelih lisica sa srebrno-crnim lisicama, međutim umjesto bijelih trebali bi se pojaviti platinasti, što se lako može provjeriti ispisivanjem odgovarajućih križeva.
Podjela dobivena u testnom križanju - pojava bjelopute i gruzijske bijele lisice - može se objasniti ili interakcijom dva (ili tri) blisko povezana gena ili interakcijom tri alela s monogenskom razlikom između izvornih oblicima.

Naglo smanjenje održivosti bijelih lisica, primijećeno u eksperimentu, govori u prilog djelovanju alelnih mutacija, budući da su u ovom slučaju bijele lisice spojevi genotipa dviju dominantnih mutacija jednog gena, obje s recesivnim letalnim učinkom. Teško je očekivati smanjenje vitalnosti kada dva različita gena međusobno djeluju (komplementarnost). Stoga su zaključili da su bijela, bjeloputa, gruzijska bijela, platinasta i srebrno-crna boja kod lisica kontrolirane nizom alela jednog gena, od kojih su tri dominantna s recesivnim letalnim učinkom. Genotipovi lisica: bijela A 1 / A 2, A 1 / A 3, A 2 / A 3; bjelo lice A 1/a; platina A 3/a; gruzijska bijela A 2/a; srebrno-crna a/a (prema Belyaev i sur., 1973).
Treba naglasiti da s višestrukim alelizmom u slučaju heterozigotnosti izvornih oblika, maksimalni broj fenotipskih klasa u podjeli ne može biti 3, već 4, kao u gore opisanom slučaju. U populaciji se broj mogućih genotipova s višestrukim alelizmom višestruko povećava; može se odrediti formulom: 1 / 2n (n+1), gdje je n broj alela. Na primjer, ako postoji 7 alela za lokus, broj mogućih genotipova u populaciji bit će 28: 1 / 2 × 7 × 8 = 28.
Učinak recesivnih letalnih mutacija na segregaciju
Recesivna autosomna i spolno vezana mušica, uzrokujući smrt homozigota u križanjima heterozigota za letal, mogu utjecati na cijepanje gena povezanih s letalom. U ovom slučaju, omjer potomaka u cijepanju ovisi o udaljenosti između gena koji se proučava i letala, kao i o tipu heterozigota – u cis- ili trans-položaju geni se unose u heterozigot. Za identifikaciju smrtonosnih, obično se izvode razna testna križanja. Primjer analize letenja je zadatak br. III.4.
Problem br. III.4
U liniji Drosophila iz linije br. 100, koja sadrži inverzije, polovica ženki imala je sivu, polovica žutu boju tijela, a pokazalo se da su žute ženke sterilne. Svi mužjaci ove linije bili su žute boje. Omjer ženki i muškaraca bio je drugačiji od normalnog, više poput podjele 2♀♀:1♂♂. Pretpostavlja se da je nedostatak mužjaka u liniji br. 100 vjerojatno posljedica prisutnosti sivih ženki na X kromosomu, koje su očito heterozigotne - linija se dijeli na sive i žute ženke. Kako bi se utvrdila genetska struktura ove linije i testirala pretpostavka o prisutnosti letala u X kromosomu ženki, obavljena su križanja čiji su rezultati prikazani u nastavku.
Uzajamni križevi

Sive ženke iz F 1 bili su pojedinačno križana sa žutim mužjacima iz linije br. 100.

Analiza
Na temelju analize napisat ćemo sheme svih križanja.

Predložene hipoteze prilično dobro objašnjavaju sve rezultate. No ostaje nejasno pitanje razloga sterilnosti žutih ženki u retku br.
U bilje mnoge recesivne mutacije povezane su s nedostatkom ili odsutnošću klorofila, što dovodi ili do smanjenja održivosti biljaka ili do njihove smrti u različitim fazama razvoja. To uzrokuje odstupanja u cijepanju, a također čini nužnim uzeti u obzir cijepanje ne samo u sadnicama, već iu kasnijim fazama razvoja kako bi se odredio udio smrti biljke i priroda nasljeđivanja svojstva. Dakle, kukuruz je homozigotan za gen wd (nedostatak bijele boje) imati bijele presadnice (podijeliti na presadnice 3/4 zelene : 1/4 bijele). Međutim, nakon 1-3 tjedna, sve bijele biljke odumiru nakon što potroše rezerve hranjivih tvari sjemena, au najnaprednijim fazama razvoja biljke, rascjep nestaje - 3 zelene: 0 bijele. Slične mutacije poznate su u grašku, ječmu, raži, pšenici itd.
Ostale mutacije uzrokuju smrt samo dijela jedinki u određenom stupnju razvoja, što dovodi do smanjenja udjela recesivnih u rascjepu i promjene u omjeru fenotipova - 4:1, 5:1 itd. Ovi omjeri variraju, budući da, u pravilu, održivost takvih mutanata uvelike ovisi o uvjetima.
Kod ljudi smanjena vitalnost i letalitet zbog djelovanja recesivnih mutacija očituje se u različitim razdobljima embriogeneze i u različitim fazama razvoja. Razlozi smanjenja sposobnosti preživljavanja i smrtonosnog učinka mogu se povezati i s mutacijama gena i s kromosomskim abnormalnostima. Citogenetička analiza pobačenih embrija omogućuje nam utvrđivanje uzroka smrti mnogih od njih. U prosjeku se zbog kromosomskih aberacija u svim fazama trudnoće događa više od 42% spontanih pobačaja: značajan udio novorođenčadi s kromosomskim aberacijama umire tijekom prve i narednih godina života.
Među smrtonosnim genskim mutacijama koje dovode do fetalne smrti ili smrti u djetinjstvu mogu se navesti recesivne mutacije koje uzrokuju talasemiju, anemiju srpastih stanica, cističnu fibrozu, kongenitalnu ihtiozu, anencefaliju (odsutnost mozga), fenilketonuriju itd.
Za proučavanje smrtonosnih mutacija ili mutacija koje smanjuju vitalnost kod ljudi, naširoko se koriste citogenetske i biokemijske metode analize, proučavanje strukture i aktivnosti enzima u zdravlju i bolesti, kao i kod heterozigotnih nositelja; kromatografija, različite vrste elektroforeze.
Letalni geni su mutacijski geni koji uzrokuju smrt jedinke prije nego što ona dostigne spolnu zrelost. Mogu biti dominantni, recesivni ili vezani za spol. Obično pokazuju svoj učinak u homozigotnom stanju, au heterozigotnom smanjuju vitalnost. Penetracija je sposobnost gena da se fenotipski manifestira, izražava se u % i može biti potpuna ili nepotpuna. Potpuna – kod svih jedinki populacije koje imaju određeni gen, manifestira se kao osobina. Nepotpuno - neki pojedinci imaju gen, ali se ne manifestiraju prema van. Ekspresivnost je stupanj izraženosti neke osobine, tj. ista je osobina izražena različitim intenzitetom kod različitih pojedinaca.
U prisutnosti različitih smrtonosnih gena, organizmi umiru u različitim fazama razvoja. U pravilu, letalni učinak takvih gena je recesivan, tj. očituje se samo kada su u homozigotnom stanju. Kada se pojave mutacije s dominantnim letalnim učinkom, organizam umire bez stvaranja potomstva.
Međutim, bilo je slučajeva gdje su smrtonosni geni, uzrokujući vidljive promjene u heterozigotnom stanju, čak postali korisni s ekonomske točke gledišta. Dakle, među ovacama karakulske pasmine postoje životinje s lijepom srebrno-sivom bojom kože, koja se cijeni skuplje od običnog crnog karakula.Kod križanja sivih ovaca sa sivim ovnovima pokazalo se da su uvijek heterozigotni. Proučavajući uzroke ove pojave, utvrđeno je da je među sivim janjadima dobivenim takvim križanjem otprilike 1/3, ili oko 25% ukupnog potomstva, oboljelo od kroničnog bubnjića i uginulo. Uzrok bolesti su poremećaji aktivnosti parasimpatičkog živčanog sustava. Križanjem sivih ovnova s crnim ovcama ili sivih ovaca s crnim ovnovima potomstvo je dalo 50% sivih i 50% crnih janjadi, a siva janjad nije obolijevala. Pokazalo se da je u homozigotnom stanju gen koji je uzrokovao razvoj sive boje imao recesivni letalni učinak.
Lisice imaju dragocjenu boju krzna - platinastu, uzrokovanu dominantnim genom koji u homozigotnom stanju uzrokuje smrt embrija u ranoj fazi razvoja. U takvim slučajevima dolazi do promjene omjera fenotipova u drugoj generaciji, jer od tri dominantna oblika jedan (homozigot za letalni gen) umire, čime omjer fenotipova postaje 2:1.
Slično cijepanje zabilježeno je i kod zrcalnog šarana, kod kojeg je uočeno nasljeđivanje nerazvijenosti ljuski: sačuvana je samo na središnjoj liniji tijela, ostatak je bio gol, zbog čega se naziva linearnom. Kada se linearni šarani međusobno križaju, potomci se uvijek dijele u omjeru 2 linearna: 1 s normalnim ljuskama, jer embriji homozigotni za gen koji uzrokuje nerazvijenost ljuski umiru u ranim fazama razvoja.
U kokoši homozigotnih za alel koji uzrokuje uvijanje perja, nepotpuni razvoj perja ima nekoliko fenotipskih učinaka. Ovi pilići nemaju dovoljnu toplinsku izolaciju i pate od hlađenja. Kako bi nadoknadili gubitak topline, razvijaju niz strukturnih i fizioloških prilagodbi, ali te su prilagodbe neučinkovite i smrtnost među takvim kokošima je visoka.
Učinak smrtonosnog gena jasno se vidi u nasljeđivanju boje dlake kod miševa. Divlji miševi obično imaju sivo krzno, poput agoutija; ali neki miševi imaju žuto krzno. Križanjem žutih miševa dobiju se i žuti miševi i agouti u omjeru 2:1. Jedino moguće objašnjenje za ove rezultate je da je žuta boja dlake dominantna kod agoutisa i da su svi žuti miševi heterozigoti. Atipični Mendelov odnos objašnjava se smrću homozigotnih žutih miševa prije rođenja. Nekropsije gravidnih žutih miševa križanih sa žutim miševima otkrile su mrtve žute mladunce u njihovim maternicama. Ako bi se križali žuti miševi i agouti, tada ne bi bilo mrtvih žutih miševa u maternicama trudnih ženki, jer takvim križanjem ne može biti potomaka homozigotnog za gen žute vune.
Promjena fenotipske segregacije u omjeru 3:1 u drugoj generaciji monohibridnog križanja povezana je s različitom sposobnošću preživljavanja F2 zigota. Različita sposobnost preživljavanja zigota može biti posljedica prisutnosti letalnih gena. Letalni je gen koji uzrokuje poremećaje u razvoju organizma, što dovodi do njegove smrti ili deformacije.
Proučavanje kongenitalnih anomalija pokazalo je da je s različitim letalnim genima smrt jedinki različita i može se dogoditi u različitim fazama razvoja.
Prema klasifikaciji koju je predložio Rosenbauer (1969), geni koji uzrokuju smrt 100% jedinki prije nego što dostignu zrelost nazivaju se letalnim, više od 50% nazivaju se subletalnim (poluletalnim), a manje od 50% nazivaju se subvitalnim. Međutim, treba napomenuti da je ova podjela u određenoj mjeri proizvoljna i ponekad nema jasne granice. Primjer je golotinja povezana sa spolom kod kokoši. Gotovo polovica golih pilića ugine u zadnja 2-3 dana inkubacije. Od izleženih pilića, otprilike polovica ugine prije 6. tjedna starosti ako se uzgajaju na temperaturi od 32-35 °C. Ali ako se temperatura u grijaćim pilećima poveća za 5,5 °C, tada će uginuti znatno manje golih pilića. 4-- Nakon 5 mjeseci, goli pilići dobiju rijetko perje i već su sposobni izdržati prilično niske temperature.
U prirodnim uvjetima ova će mutacija vjerojatno biti smrtonosna i dovesti do 100% smrtnosti ptica. Gornji primjer pokazuje da priroda manifestacije polu-letalnog gena može uvelike ovisiti o uvjetima okoline.
Letalni geni mogu biti dominantni ili recesivni. Među prvim smrtonosnim čimbenicima otkriven je alel koji je uzrokovao žutu obojenost miševa. Dominantan je gen žute boje (Y). Njegov recesivni alel (y) u homozigotnom stanju uzrokuje pojavu crne obojenosti. Međusobno križanje žutih miševa dalo je dva dijela žutih miševa i jedan dio crnih, tj. rezultat je bio podijeljen u omjeru 2:1, a ne 3:1, kako proizlazi iz Mendelovog pravila. Ispostavilo se da su svi odrasli miševi heterozigoti (Yy). Prilikom međusobnog križanja trebali su proizvesti jedan dio homozigotnog potomstva za žutu boju (IT), ali ono umire u embrionalnom razdoblju, dva dijela heterozigota (Yy) bit će žuta, a jedan dio homozigota za recesivna osobina (yy) bit će crna. Shema križanja izgleda ovako:
Na isti način, siva boja dlake nasljeđuje se kod karakul ovaca (Sokolskie, Malich itd.), platinasta boja kod lisica, distribucija ljuski kod linearnih šarana itd.
Smrtonosni geni su u većini slučajeva recesivni i stoga mogu dugo ostati latentni.
Životinja koja je potpuno zdrava i normalnog fenotipa može biti nositelj letalnog gena, čiji se učinak otkriva tek nakon prijelaza u homozigotno stanje. Letalni geni najčešće prelaze u homozigotno stanje tijekom inbridinga. U praksi stočarstva pri uzgoju konja zabilježen je slučaj uginuća 25 ždrebadi 2.-4. dana nakon rođenja od deformacije rektuma - odsutnosti anusa (Atresia ani). Ispostavilo se da su svi pastusi i kobile koji su rodili takvu abnormalnu ždrebad potekli od istog pastuha. Bio je heterozigot za smrtonosni gen (LI). U početku je ovaj pastuh, kada je križan s normalnim kobilama (LL), dao potomke koji su bili normalni u fenotipu, ali u smislu genotipa, polovica potomaka bila je normalna (LL), a polovica su bili heterozigoti (LI), noseći recesivna sklonost (0 letalni gen. U srodstvu heterozigotnih životinja (Y x Y) pojavila su se ždrijebeta homozigotna za letalni gen (II) s deformitetom rektuma. Sva su uginula.
Poznati su deseci anomalija kod domaćih životinja čija je pojava povezana s recesivnim ili dominantnim mutacijama gena. Ove se anomalije javljaju u pojedinim populacijama s različitom učestalošću, što ovisi o brzini mutacijskog procesa, sustavu uzgoja životinja i sl. Poznavanje specifičnih oblika kongenitalnih anomalija u životinja svake vrste, kao i učestalosti njihovog očitovanja u pojedine pasmine, nužna je veterinarima za selektivnu prevenciju širenja genetske patologije.
Anomalije kod goveda. Biološke karakteristike ove vrste životinja su kratka dlaka i relativno kasno sazrijevanje. Krava obično okoti jedno tele koje spolnu i fiziološku zrelost dostiže tek za 1,5 godinu, pa je razdoblje između prvog teljenja majke i kćeri prosječno 5 godina. Kao rezultat toga, pojava abnormalnog potomstva u stadu može značajno smanjiti razinu reprodukcije i intenzitet uzgojne selekcije stoke. U goveda je proučavan širok raspon kongenitalne šamalije, određene letalnim, polu-letalnim i subvitalnim genima. 46 anomalija uključeno je u Međunarodni popis smrtonosnih defekata pod oznakom A (tablica 43). Relativna učestalost pojedinih vrsta abnormalnosti u svakoj pasmini ili populaciji može varirati. Kod kostromske pasmine, prema našim podacima, najčešće zabilježena genetska anomalija glave je skraćenje čeljusti (tablica 44), kod jaroslavske pasmine - sindaktilija, kod holmogorske pasmine - mišićne kontrakture, kod crno-crne pasmine. Bijela - pupčana kila. Abnormalnosti središnjeg živčanog sustava bile su najčešće (21%) kod goveda u Njemačkoj.
Drugo mjesto po učestalosti registracije (14%) zauzela je složena anomalija - kombinacija pupčane kile s rascjepom trbuha i fetusa u cjelini. Učestalost anomalija, odnosno postotak abnormalnog potomstva u ukupnom broju, unutar pojedinih populacija također može biti vrlo različit i, prema prosječnim procjenama, ne prelazi 1%. Međutim, ovaj pokazatelj ovisi o cjelovitosti i točnosti snimanja anomalija. Tako su u Njemačkoj nakon organiziranja čistog računovodstva zaključili da se učestalost anomalija višestruko povećala. Pitanje je: jesu li sve anomalije podložne vizualnom promatranju? Očito ne sve. Tako je u kostromskoj pasmini prosječna učestalost svih oblika anomalija u razdoblju od 12 godina iznosila 1,15%. Učestalost ukupnog mortaliteta legla (abortirana, mrtvorođena, abnormalna, mrtva telad bez vidljivih nedostataka) u ovoj farmi bila je 10,2%. Određeni dio ove smrtnosti povezan je i s mutacijama gena koje uzrokuju ne morfološke nedostatke, već metaboličke poremećaje i druge anomalije, čije je identificiranje moguće samo posebnim metodama.
Proizvođači mogu igrati posebnu ulogu u širenju genetskih abnormalnosti kod goveda i drugih životinjskih vrsta. Od svakog oka, umjetnom oplodnjom, mogu se godišnje dobiti stotine i tisuće potomaka. Tako je od jednog bika u inozemstvu dobiveno 100 tisuća teladi. Ako se pokaže da je takav otac nositelj mutacije gena, ona će se brzo proširiti kroz pasminu. Evo nekoliko primjera iz brojnih činjenica opisanih u literaturi. Kao rezultat intenzivnog korištenja bika Prince Adolfa, dovedenog u Švedsku, i naknadnog spontanog parenja na njemu, učestalost bezdlake u nekim švedskim stadima bila je preko 5%. Ista situacija nastala je u Švedskoj nakon uvoza bika Gallus, za kojeg se pokazalo da je heterozigotni nositelj gena koji uzrokuje nedostatak udova.
U potomstvu pojedinačnih bikova crno-bijele i šarolaske rase u SAD-u i Njemačkoj zabilježeni su slučajevi rođenja patuljaste teladi s učestalošću od 23,3 odnosno 22,2%. U bivšem U Čehoslovačkoj je pri ispitivanju potomaka 166 bikova utvrđeno da su njih 43 nositelji smrtonosnih gena. U jednog bika - nositelja dominantne anomalije "rascjep usne" - nedostatak se očitovao kod 44% bikova i 71% junica iz njegovog potomstva.
U kostromskoj pasmini analizirali smo širenje skraćivanja donje čeljusti i mopsastog izgleda preko bika Burkhana, koji je i sam imao neispravnu telad u svom potomstvu; njegovi sinovi, unuci, praunuci, ženski potomci također su dali nenormalno potomstvo (slika 57). Većina teladi dobivena je inbridingom i parenjem roditelja s normalnim fenotipom i zajedničkim pretkom. Stoga možemo zaključiti da ova anomalija ima recesivni način nasljeđivanja. Slika pokazuje da je najveći broj defektne teladi registriran u potomstvu bika Zheton 3501 (unuk bika Burkhana) kada je korišten u komercijalnoj farmi, gdje su neke od krava imale isti recesivni gen u svom genotipu.
Anomalije kod svinja. Međunarodni popis smrtonosnih defekata kod svinja uključuje 18 genetskih abnormalnosti. Glavninu njih uzrokuju autosomno recesivni geni (tablica 45). Genetske abnormalnosti mogu igrati značajnu ulogu u patologiji svinja. Pogledajmo nekoliko takvih primjera. U Španjolskoj je istraživanje na 23 449 prasadi iz 2 399 legla dobivenih od nerastova pasmina Duroc, Yorkshire, Hampshire i White Chester otkrilo, tim redom, 6,21; 6.02; 9,66; 2^62% abnormalnih legla.
Prema Olivieru (1979.), 7 genetskih abnormalnosti kože, 17 kostura, 3 očiju, 13 neuromuskularnih, 6 krvi, 6 hormonskog metabolizma, 5 probavnog sustava i 9 genitourinarnog sustava opisani su kod svinja. Glavne anomalije bile su kriptorhizam, kile, pseudohermafroditizam itd. Autor studije smatra da su te anomalije posljedica djelovanja jednog gena u različitim fazama formiranja embrija.
U Danskoj je tijekom dvogodišnjeg razdoblja proučavano 6669 mrtvih prasadi iz 2936 legla kako bi se utvrdila priroda i učestalost kongenitalnih anomalija. Razne anomalije otkrivene su kod 1,4% rođene prasadi, odnosno 6,2% ubijene prije odbijanja. U postmortem pregledu 25,9% abnormalne prasadi imalo je nerazvijene zaliske, nezatvaranje anusa, subaortalnu stenozu, ektopiju srca i druge nedostatke kardiovaskularnog sustava. U 23,4% prasadi utvrđeni su različiti razvojni poremećaji motoričkog sustava. Anomalije središnjeg živčanog sustava otkrivene su u 5,9% prasadi, uključujući bifurkirani mozak i cerebralnu hidrokelu. Prekomjerni rast rektuma, tankog crijeva ili njihov nepotpuni razvoj pronađeni su u 30% prasadi, a razne hernije i ascites - u 6,8%. Rascjep usne, rascjep nepca, rinocefalitis i druge anomalije facijalnog dijela glave nađeni su u 6,1%; hermafroditizam, rascjep uretera, hidrokela bubrega i uretre - u 1,7% prasadi. Ove anomalije su se pojavile u potomcima pojedinih bikova tijekom inbreedinga, što ukazuje na nasljednu prirodu njihove pojave.
Vrlo uvjerljive dokaze o nasljednoj prirodi kriptorhizma kod svinja dobili su Fridin i Newman. Prema njihovim podacima, u Kanadi se jednostrani i obostrani kriptorhizam godišnje opaža kod 1-2% svih nerastova koji izlaze na tržište. Autori su križali kriptorhide s njihovim majkama i punim sestrama. Potomci iz takvih križanja međusobno su pareni. Kao rezultat takve selekcije i selekcije, učestalost kriptorhizma u pokusnih životinja jorkširske pasmine porasla je u prosjeku na 42,9%, a osobito kada su korištena dva proizvođača. Prilikom ispitivanja prasadi u Sjedinjenim Državama u jednoj godini, oko 400 tisuća životinja pronađeno je sa skrotalnom hernijom.
Promatranja pokazuju da je uzrok smanjene plodnosti u nerastova često hipoplazija testisa. Učestalost ove anomalije, prema istraživačima iz Njemačke, bila je 19,6%. Za reprodukciju je ostavljeno 30 takvih nerastova
Va, svaki od njih pokrivao je od 4 do 40 matica (ukupno 439 grla), ali samo četiri od njih rodile su potomstvo. Analiza je pokazala da su patološki oblici sperme kod ovih nerastova 80-100%. Svih 30 abnormalnih životinja imalo je zajedničke pretke, što ukazuje na nasljednu prirodu hipoplazije testisa i defekata spermiogeneze.
Prisutnost kraterskih bradavica kod svinja jedan je od ozbiljnih nedostataka, budući da prasadi ne dobivaju mlijeko od njih. Prema Bavarskom institutu za stočarstvo (Njemačka), učestalost ove anomalije kod njemačkih landrasa bila je 6,6%. Kao što su primijetili P. N. Kudryavtsev i dr. (MVA), broj svinja s neaktivnim kraternim sisama povećao se posljednjih godina. Broj takvih bradavica kreće se od 1 do 8. Prasad koja dobije krater bradavice umire.
Krateritet je svojstvo uzrokovano jednim autosomno recesivnim genom. To su eksperimentalno provjerili P. N. Kudryavtsev i sur. Nakon što su prethodno identificirali nerastove i nazimice koje su nositelji anomalija (KchKch), normalne ali heterozigotne jedinke (KchKch), koje rađaju prasad s kraterom u potomstvu, i normalne homozigotne svinje (KhKch), autori su izvršili križanja između ovih skupina životinja . U prvoj varijanti križano je 27 normalnih homozigotnih majki s 15 nerastova. Svih 258 potomaka bilo je normalno. U drugoj varijanti, gdje je jedan od roditelja bio homozigot (KchKch), a drugi heterozigot (Kchkch), sva su prasad također bila normalna. U trećoj varijanti križano je 13 heterozigotnih nerastova sa 16 heterozigotnih majki. Od 168 rođenih prasadi, 39 (23,2%) imalo je krater bradavice. I konačno, u četvrtoj opciji, jedan od roditelja bio je homozigot, a drugi heterozigot. Dobili su 170 prasadi, od kojih je 86 (50,5%) imalo normalne bradavice, a 84 (49,5%) bradavice s kraterima. Rezultati ovog pokusa dokazuju recesivni način nasljeđivanja kratera bradavice u svinja.
Abnormalnosti kod ovaca. Kod ovaca je opisano oko 90 kongenitalnih anomalija. Prema Dennisu i Leipoldu, većina poznatih genetskih defekata u ovaca uzrokovana je monogenim autosomno recesivnim načinom nasljeđivanja (Tablica 46). Najčešći kraniofacijalni defekti kod ove vrste životinja su kraniofacijalni defekti, posebno agnatija, kao i zakrivljenost prednjih udova, mikroagnatija, hermafroditizam, kriptorhizam, hipospadijaza, prognatija, analna atrezija, mikrotija, entropija, tortikolis, politelija, artrogripoza. Analiza je pokazala da se 55,4% defekata odnosi na mišićno-koštani sustav, 12,7 - na probavni sustav, 9,7 - na kardiovaskularni sustav, 7,1 - na urogenitalni sustav, 6 - na središnji živčani sustav, 3,5 - na anomalije. ligamenata, 3,2 - na trbušni, 1,5% - na endokrini sustav. Iako je učestalost pojedinačnih nedostataka niska, kumulativni doprinos svih anomalija može uzrokovati štetu na uzgajalištima. U Novom Zelandu, zemlji s razvijenim uzgojem ovaca, učestalost smrtonosnih nedostataka bila je oko 1% mrtve janjadi. U SAD-u su smrtonosni nedostaci u prva 2 tjedna života uočeni kod 11,4% janjadi.
Prosječna stopa embrionalne smrtnosti ovaca je 20%. Ovo ukazuje na to da mnogi smrtonosni geni koji se ne mogu identificirati mogu biti aktivni tijekom tog razdoblja.
Individualne genetske abnormalnosti kod ovaca mogu biti raširene. Tako je u Bugarskoj u stadima Merino ovaca uočena visoka smrtnost janjadi u ranom postnatalnom razdoblju. Nastala je kao posljedica činjenice da janjad nije dobivala mlijeko zbog nedostataka u vimenu njihovih majki: od hipoplazije s malim ostacima žljezdanog tkiva do njegovog potpunog odsustva. Učestalost ove nasljedne anomalije u različitim stadima kretala se od 6 do 40%.
Kod ovaca je često uočen kriptorhidizam, koji je bio u kombinaciji s takvim sekundarnim spolnim obilježjem kao što je poledness. Ovnovi za pušenje imaju nisku plodnost. Strogom selekcijom u uzgoju uspjelo se stvoriti tip ovnova s normalnom plodnošću, no među njima se nalaze i abnormalne jedinke.
Anomalije kod ptica. Ptice, prvenstveno kokoši, najviše su proučavane u odnosu na genetiku abnormalnosti. Međunarodni popis smrtonosnih mana uključuje 45 anomalija kod kokoši, 6 kod purana i 3 kod pataka. Najčešće abnormalnosti kljuna su (papagajski kljun, prekriženi kljun). Njihova je učestalost, prema Williamu i sur., 1,1% gubitaka tijekom inkubacije jaja kokoši White Leghorn i Rhode Island. Abnormalnosti kljuna također su česte kod pataka.
Max Gibbon i Shakelfurd opisali su anomaliju pri križanju bijelih leghorna s buttercampsima i bantamima i naknadnom uzgoju "u sebi" - polidaktiliji. Osim toga, sindaktilija i pernate noge uočene su kod pilića. Učestalost sindroma bila je 16,8%. Prilikom križanja abnormalnog F2 pijetla s fenotipski normalnim kokošima uočena je podjela - pola normalnih i pola abnormalnih jedinki (1:1). Utvrđeno je da ovaj kompleks svojstava kontrolira jedan autosomni gen, koji ima poluletalni učinak, budući da je embrionalno i postembrionalno preživljavanje abnormalnih pilića vrlo nisko.
Anomalije kod konja. Od nasljednih anomalija kod konja, 10 ih je uključeno u Međunarodni popis smrtonosnih mana. Među njima su 3 anomalije kostura, 2 reproduktivnog sustava, 2 bubrega i mišića, po jedna anomalija crijeva, živčanog sustava i organa za vid.
U konja zaprežnih pasmina češća je atrezija debelog crijeva. Prevalencija ove anomalije zabilježena je u potomstvu pastuha Superba pasmine Percheron. Također je opisan u ždrebadi čistokrvne jahaće pasmine istočnoafričkog podrijetla. Nesavršena epiteliogeneza zabilježena je kod teglećih pasmina. Ataksija, nazvana Oldenburg, pronađena je u Oldenburg ždrijebadi u Njemačkoj. Proširila se u redu 9. Jedna od najčešće bilježenih anomalija kod konja je pupčana kila. Nalazi se u lakim i teškim pasminama.
Kod konja je poznato nekoliko drugih genetskih i nasljedno-okolišnih anomalija. Tako u SAD-u konji nekoliko pasmina imaju ždrijebe s neobičnom bijelom mrljom, zvanom "overo". Kod križanja konja tipa "overo" rađaju se ždrebad s ružičastom kožom, u kojoj se uočava hipoplazija crijevnog trakta i izoeritroliza, kao i kolike, što dovodi do smrti.
U Engleskoj su životinje s poremećenom koordinacijom pokreta - "bolest kolebanja" - registrirane i proučavane kod konja polukrvne jahaće pasmine. Utvrđena je genetska predispozicija za ovu anomaliju.
Dokazana je nasljednost dermatoza udova u konja. Defekti s nasljednom predispozicijom su česte kronične deformirajuće upale skočnog zgloba, takozvane „štake” u ždrebadi, kao i kronične aseptičke upale kruničnog bloka kopita, uočene uglavnom kod trkaćih konja i trkaćih konja. konji.









- Pokretnine stečene nakon 01
- Porez na imovinu stečenu nakon 01
- Morski konjic - izvješće o poruci Koliko dugo morski konjic postoji na zemlji
- Vanilin kolačići s mliječnom čokoladom
- Recept za palačinke s kiselim kvascem s fotografijom
- Povijest izuma votke
- Nasljeđe svetih otaca: pravoslavne parabole o životu i moralu i biblijski citati
- Kraljevsko meso, novogodišnji recept za kuhanje kraljevskog mesa Kraljevsko meso s krumpirom u pećnici
- Bijeli kvas za okrošku Napravite kvas od okroške kod kuće
- Kemija (građa atoma) (prezentacija)
- Tradicije i običaji Engleske koji će vas iznenaditi
- Nasljedstvo i razvoj Je li naslijeđe toliko važno?
- Harry Potter knjige na engleskom
- Organizacijski čimbenici koji uzrokuju stres
- Recept za pirjani kupus u laganom kuhalu: jednostavan, brz i ukusan
- Osmica štapova, opis i karakteristike karata
- Odrezak i medaljoni od lososa
- Sažetak individualnog logopedskog sata o automatizaciji zvuka Sažetak automatizacije zvuka l u rečenicama
- Nižnji Novgorod Državno pedagoško sveučilište nazvano po
- Svi oni koji su razapeli Krista umrli su strašnom smrću.Je li Poncije Pilat doista bio tamo?