Рецесивни, доминантни, летални гени, мутации. III.2.8
Досега разглеждахме човешката генетика според видовете наследство; сега ще се обърнем към една много важна категория гени, а именно леталните и сублеталните гени. Леталните и сублеталните гени са клас мутации с подчертано изразени отрицателни ефекти върху развитието. Както ще видим по-късно, мутациите в много случаи влошават и унищожават наследствеността на човека. Това


влошаването от мутации е особено изразено под влиянието на летални гени.
Леталните и сублеталните алели могат да бъдат доминиращи или рецесивни. При доминиращ леталитет детето умира в пренаталния период или в ранна детска възраст. Съвсем ясно е, че такива гени не могат да бъдат наследени. Тяхното присъствие показва появата на мутации. Общата честота на такива доминиращи смъртоносни случаи не е толкова малка. Техният брой е особено голям под въздействието на радиация. В генетиката на мишките и Drosophila доминантните летали се използват широко, когато се анализира зависимостта на честотата на мутациите от дозата и когато се анализират други характеристики на мутагенните ефекти. При наличието на сублетални, иначе наречени полулетални гени, детето умира в ранна детска възраст или малко по-късно, но като правило преди да достигне пубертета. Пример за доминантна сублетална мутация е епилоята, заболяване, характеризиращо се с ефекта на цял синдром от негативни симптоми. Децата с епилоия имат патологични израстъци на кожата, характеризират се с умствена изостаналост, епилепсия, тумори на сърцето, бъбреците и други органи. В редки случаи пациентите с епилоя оцеляват и се женят за здрав съпруг. Половината от децата от такъв брак се оказват болни от вродена епилоя (фиг. 131).
Друг ярък пример за сублетална доминантна мутация е заболяването ретинобластом. Това


доминиращият ген причинява раков тумор в окото, което почти винаги води до смърт на детето в ранна възраст.
По-голямата част от вродените деформации обаче се падат на рецесивните летални и сублетални. В този случай рецесивните вредни гени са скрити в генотипа на привидно здрави родители, но когато две такива хетерозиготи се женят, около 25% от децата им се оказват болни.
Фигура 132 показва основните видове кръстосвания в човешките популации, които се извършват при наследяване на рецесивни летални и сублетални гени. На квадрат 1 представя типа бракове между генетично нормални хора. При двамата родители и всички техни потомци всички алели са представени от нормални гени (алел +). На квадрат 2 изобразява брак между генетично здрав човек и хетерозигота (символ a). В този случай отрицателният алел се предава на половината от потомството, но остава скрит. На квадрат 3 генетично здрав човек се жени за болен (символ аа),всички деца са видимо здрави, но в хетерозиготно състояние носят сублетална мутация А.На квадрат 4 хетерозигот се жени за пациент; половината от децата им са видимо здрави, но носят сублетална мутация, другата половина са болни. Накрая квадрат 5 в случай на брак на двама болни (символи аа)цялото потомство ще бъде болно. Вродените заболявания, причинени от действието на рецесивни летални и сублетални гени, включват детска амавротична идиотия, при която мозъчната тъкан се дегенерира, настъпва слепота и детето умира; вродена кожна патология с наличие
дълбоки кървящи пукнатини - ихтиоза (фиг. 133); вродена детска парализа и редица други вродени заболявания.
Много ранни спонтанни аборти са причинени от изтичане на летални генотипове.
Известни са доминантни мутации, които при хетерозиготите определят някои повече или по-малко неутрални черти, които обаче имат летален ефект в хомозиготно състояние. Например алелът на пигментацията на ксеродерма в хетерозиготно състояние причинява тежки лунички. Хомозиготите обаче развиват пигментна ксеродерма. Това е сублетално заболяване (фиг. 134), при което действието на светлината причинява увреждане на кожата, което след това води до злокачествен растеж.
Един от алелите при човек в хетерозиготно състояние причинява излишък на холестерол в кръвта. Въпреки това, децата, хомозиготни за този алел, развиват тежка сублетална аномалия.
Във всички тези случаи имаме класическа картина на унаследяване на доминанти с рецесивен летален ефект, установен преди това в експерименти с дрозофила, мишки и други организми. Например, мишката има добре познат алел A Y, което кара кожата да пожълтява при хетерозиготите. Въпреки това всички опити да се получи чиста раса жълти мишки останаха неуспешни. При кръстосването на две жълти мишки средно две жълти мишки винаги произвеждат една с различен цвят, например черна. Естеството на това явление се обяснява с факта, че генът за доминиращия жълт цвят се оказва
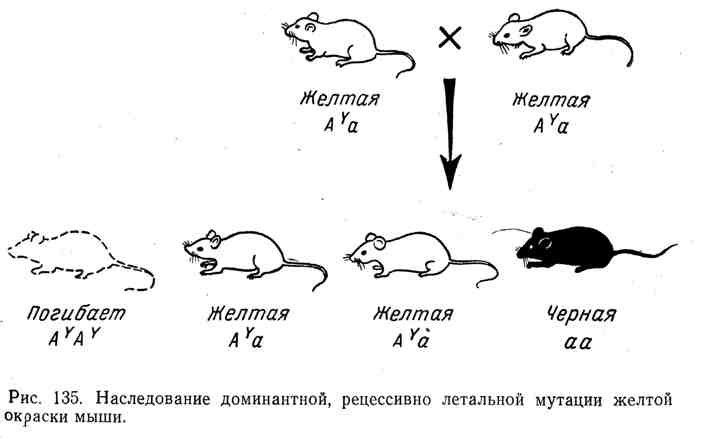
рецесивен летален. Всички хомозиготи( A Y A Y) починал. Жълтата мишка винаги е хетерозиготна, заедно с алелаA Y той съдържа друг алел, напр. А(черен цвят). Ходът на наследяване при кръстосване на хетерозиготи A Y a и разделяне по формулата 2: 1 са показани на фигура 135.
- Източник-
Дубинин, Н.П. Хоризонти на генетиката / Н.П. Дубинин. – М.: Образование, 1970.- 560 с.
Преглеждания на публикация: 121
Нееднаквата жизнеспособност на зиготите от различни генотипове може да бъде свързана с доминантни или рецесивни летални мутации, които възникват в гаметите на родителските индивиди. Те могат да се реализират на различни етапи от ембриогенезата или в постембрионалния период. Естествено, смъртта на някои зиготи засяга естеството на разцепването.
Влияние върху разделянето на доминантни гени с рецесивен летален ефект
Доминантните гени с рецесивен летален ефект имат плейотропен ефект;от една страна, те определят доминиращия характер на проявлението на всяка черта в хетерозигота, от друга, те причиняват смъртта на индивиди в хомозигота, т.е. проявяват леталния си ефект като рецесивен. Такива гени са известни при много животни - жълто оцветяване при мишки, къси крака при пилета, линейно лющене при шаран, платинено оцветяване при лисици, ширази оцветяване при овце и много други. Те предизвикват отклонение от цепителността 3:1, превръщайки я в цепителност 2:1. Анализът на сегрегациите, включващи доминантни гени с рецесивен летален ефект, се усложнява от факта, че в сравнително малки проби не винаги е възможно да се направи разлика между разделянето 3:1 и 2:1, а статистическият метод на тестване с помощта на критерия χ 2 не позволява да се направи избор от хипотези. Например, ако при кръстосване на жълти мишки една с друга се получи разделяне 65 жълти: 32 черни, методът %2 не отхвърля две хипотези: 1 - разделянето в експеримента съответства на разделяне 3:1, χ 2 = 3.29, p>0.05; 2 - разделянето в експеримента съответства на разделянето 2:1, χ 2 = 0.17, p>0.05.
Само при големи проби разделянето 2:1 се различава от разделянето 3:1.. При обобщаване на данните за наследяването на жълт цвят при мишки, получени от различни изследователи, разделянето при кръстосване на жълти мишки една с друга е 2386 жълти: 1235 черни - ∑ = 3621, χ 2 = 0,96 при H 0 - 2: 1 ( при H 0 - 3:1 в този случай χ 2 = 160!).
Очевидно е, че различни методи за генетично изследване, например провеждане на аналитични и обратни кръстосвания, могат да допринесат за правилното решаване на проблема с наследството. В примера, обсъден по-горе, това са кръстове: жълто от F 1 × черно; жълто от F 1 × жълто P. В първия случай трябва да има разделяне 1:1, във втория - отново 2:1.
По-трудно е да се установи, че има отклонение и то се дължи на различна жизнеспособност на зиготите, когато смъртта на някои зиготи настъпва в постембрионалния стадий. В тази връзка е необходимо да се следи плодовитостта и смъртността в потомството, ако има предположение за леталност. Основата за предположението за смъртност е невъзможността да се получи еднородно потомство при размножаване на индивиди от определен фенотип.Примери за анализ са дадени в задачи № III.2 и III.3.
Проблем № III.2
В потомството на кръстосване на сребърно-самурови норки помежду си в F 1 винаги се наблюдава разделяне: появяват се сребърно-самурови и кафяви индивиди.
В един експеримент, няколко кучила кръстоски сребърно-самур са дали 93 сребристо-самурови и 43 кафяви кученца, със среден размер на котилото от 3,66 кученца. Чрез кръстосване на сребрист самур с кафяви норки са получени 39 кафяви и 41 сребристо самур. При кръстосване на кафяви норки помежду си се получава само кафяво I потомство. Размерът на котилото при последните две кръстоски беше 4,9-5,2 кученца.
Обяснете разделянето, определете генотиповете на сребърно-самур и кафяви норки.
Анализ
Тъй като сребристите самурови норки винаги произвеждат сегрегация, когато се кръстосват една с друга и с кафяви, те очевидно са хетерозиготни. За да определите броя на гените, помислете за разделянията в експеримента:
H 0 - разлики в един ген, разделяне 3:1, χ 2 = 3.2, p>0.05. Отклонението е случайно, хипотезата не се отхвърля.

Съотношението съответства добре на разделяне 1:1 за един ген, χ 2 = 0.05, p>0.80. Отклонението е случайно, хипотезата не се отхвърля.
Сравнението на данните за плодовитостта на норките при различни кръстове показва частична смърт на зиготите при кръстосване на сребърно-самурови норки. Можем да предположим смъртта на доминантните хомозиготи. Тогава генотипът на сребристо-самулевите норки е Аа, на кафявите - аа, а разделянето при кръстосването не е 3:1, а 2:1 (χ 2 = 0,17, p>0,05). За окончателното потвърждение на тази хипотеза е необходимо да се извърши допълнително кръстосване между индивиди от сребърен самур, за да се увеличи извадката и да се тества статистически хипотезата 2:1. В някои случаи е необходимо да се извърши по-сложен анализ.
Проблем № III.3
При кръстосването на платинени, белолики и грузински бели лисици със сребристо-черни е установено, че платинените, белоликите и грузинските бели цветове, които причиняват общо отслабване на пигментацията и появата на различни пигменти, не са полови. -свързани. Кръстосването на всеки от тези мутанти със сребристо-черни води до сегрегация 2:1, като сребристо-черният цвят е рецесивен. Следователно всеки от тях се контролира от един доминантен ген с рецесивен летален ефект. Смъртта на някои мутанти се доказва от данни за размера на котилото: сребристо-черните имат 4,5 кученца на котило, белите 3,5, платинените и грузинските бели - по-малко от 3,5.
За да се установи дали тези мутации засягат един или различни гени, бяха извършени кръстосвания, резултатите от които са дадени по-долу. Тестът за алелизъм не е приложим в този случай, тъй като мутациите са доминиращи:

Обяснете разделянето, определете генотипа на всички форми.
Анализ
1. Разцепванията в пресичане 1 и 2 съответстват на съотношението 1:1:1:1 (χ 2 = 5,83, p>0,10 за пресичане 1, χ 2 = 0,55, p>0,90 за пресичане 2). Те могат да бъдат резултат от няколко причини.
1. Тъй като при кръстоски 1 и 2 има 4 класа на разцепване и съотношение 1:1:1:1, може да се приеме, че изследваните форми се различават по два независимо унаследени доминантни гена с рецесивен летален ефект, които взаимодействат по типът на допълване. В този случай един от гените е представен от два различни доминантни алела.

2. Сегрегацията 1:1:1:1 може да е следствие от тясната връзка на тези два гена при липса на кръстосване между тях:

(В решетките на Punnett са дадени фенотипни радикали - гени, които проявяват своя ефект във фенотипа.)
3. Разликите в цвета са причинени от три независимо наследени доминантни гена с рецесивен летален ефект. При първото (а) и второто (б) кръстосване, разделянето се случва в два различни гена.

4. Сегрегацията 1:1:1:1 може да е резултат от тясната връзка на три взаимодействащи гена при липса на кросинговър.

* (Редът на гените може да е различен, даден е произволно.)
5. Сегрегацията в съотношение 1:1:1:1 може да бъде резултат от различия в един ген, представен от поредица от четири алела, три от които са доминиращи с рецесивен летален ефект, а четвъртият е рецесивен:

За да се направи избор между тези хипотези, беше направен кръст - бели със сребристо-черни.
Ако една черта се контролира от два или три гена, тогава с независимото им наследяване може да се очаква появата на четири фенотипни класа:

Подобен резултат трябва да има при кръстосване на бели, получени от платинени или грузински бели лисици със сребристо-черни, но вместо бели трябва да се появят платинени, което може лесно да се провери чрез изписване на съответните кръстове.
Разделянето, получено при тестовото кръстосване - появата на белолици и грузински бели лисици - може да се обясни или с взаимодействието на два (или три) тясно свързани гена, или с взаимодействието на три алела с моногенна разлика между оригинала форми.

Рязкото намаляване на жизнеспособността на белите лисици, отбелязано в експеримента, говори в полза на действието на алелни мутации, тъй като в този случай белите лисици са генотипни съединения на две доминантни мутации на един ген, и двете с рецесивен летален ефект.Трудно е да се очаква намаляване на жизнеспособността, когато два различни гена взаимодействат (комплементарност). Следователно те стигнаха до заключението, че белият, белоликият, грузинският бял, платиненият и сребристо-черният цвят при лисиците се контролират от поредица от алели на един ген, три от които са доминиращи с рецесивен летален ефект. Генотипове на лисица: бяло A 1 / A 2, A 1 / A 3, A 2 / A 3; бяло лице A 1/a; платина А 3/а; грузинско бяло A 2/a; сребристо-черен а/а (по Беляев и др., 1973).
Трябва да се подчертае, че с множествен алелизъм в случай на хетерозиготност на оригиналните форми, максималният брой фенотипни класове в разделянето може да не е 3, а 4,както в описания по-горе случай. В една популация броят на възможните генотипове с множествен алелизъм се увеличава многократно; може да се определи по формулата: 1 / 2n (n+1), където n е броят на алелите. Например, ако има 7 алела за локус, броят на възможните генотипове в популацията ще бъде 28: 1 / 2 × 7 × 8 = 28.
Ефект на рецесивни летални мутации върху сегрегацията
Рецесивна автозомна и свързана с пола муха,причинявайки смъртта на хомозиготи при кръстосвания на хетерозиготи за летален, те могат да повлияят на разделянето на гени, свързани с летален.В този случай съотношението на потомците в разделянето зависи от разстоянието между гена, който се изследва и леталния, както и от вида на хетерозиготата - в цис- или транс-позиция гените се въвеждат в хетерозиготата. За да се идентифицират леталните, обикновено се извършват различни тестови кръстосвания. Пример за летателен анализ е задача № III.4.
Проблем № III.4
В линията Drosophila от линия № 100, съдържаща инверсии, половината женски имат сив, половината жълт цвят на тялото, а жълтите женски се оказват стерилни. Всички мъже от тази линия бяха жълти на цвят. Съотношението между жени и мъже беше различно от нормалното, по-скоро като 2♀♀:1♂♂. Предполага се, че липсата на мъжки в линия № 100 вероятно се дължи на наличието на сиви женски на X хромозомата, които очевидно са хетерозиготни - линията се разделя на сиви и жълти женски. За да се установи генетичната структура на тази линия и да се тества предположението за наличието на смъртоносни в X хромозомата на женските, бяха извършени кръстове, резултатите от които са представени по-долу.
Реципрочни кръстове

Сиви женскиот F 1 бяха индивидуалнокръстосани с жълти мъжки от линия No100.

Анализ
Въз основа на анализа ще напишем схемите на всички кръстовища.

Предложените хипотези обясняват всички резултати доста добре. Неизяснен обаче остава въпросът за причината за стерилитета на жълтите женски в ред № 100. Каним читателя да помисли върху този въпрос и да предложи хипотеза, която да го обясни.
U растениямного рецесивни мутации са свързани с липса или липса на хлорофил, което води или до намаляване на жизнеспособността на растенията, или до тяхната смърт на различни етапи от развитието. Това причинява отклонения в разделянето и също така налага да се вземе предвид разделянето не само в разсада, но и в по-късните етапи на развитие, за да се определи делът на смъртта на растенията и естеството на наследяване на признака. Така царевицата е хомозиготна за гена wd (дефицит на бяло)има бели разсад (разделени на разсад 3/4 зелени: 1/4 бели). Въпреки това, след 1-3 седмици, всички бели растения умират след изразходване на хранителните запаси от семената и в най-напредналите стадии на развитие на растенията, разделянето изчезва - 3 зелено: 0 бяло. Подобни мутации са известни при грах, ечемик, ръж, пшеница и др.
Други мутации причиняват смъртта само на част от индивидите на определен етап от развитието, което води до намаляване на дела на рецесивите в разцепването и промяна в съотношението на фенотипите - 4:1, 5:1 и т.н. Тези съотношения варират, тъй като по правило жизнеспособността на такива мутанти зависи до голяма степен от условията.
При хоратанамалена жизнеспособност и леталност поради действието на рецесивни мутации се проявява в различни периоди на ембриогенезата и на различни етапи от развитието. Причините за намаляването на жизнеспособността и леталния ефект могат да бъдат свързани както с генни мутации, така и с хромозомни аномалии. Цитогенетичният анализ на абортирани ембриони ни позволява да определим причината за смъртта на много от тях. Средно поради хромозомни аберации на всички етапи от бременността се случват повече от 42% от спонтанните аборти: значителна част от новородените с хромозомни аберации умират през първата и следващите години от живота.
Сред смъртоносните генни мутации, които водят до смърт на плода или смърт в ранна детска възраст, могат да се назоват рецесивни мутации, които причиняват таласемия, сърповидно-клетъчна анемия, кистозна фиброза, вродена ихтиоза, аненцефалия (отсъствие на мозък), фенилкетонурия и др.
За изследване на смъртоносни или намаляващи жизнеспособността мутации при хора се използват широко цитогенетични и биохимични методи за анализ, изучаващи структурата и активността на ензимите в здравето и болестта, както и при хетерозиготни носители; хроматография, различни видове електрофореза.
Смъртоносните гени са мутационни гени, които причиняват смъртта на индивида преди да достигне полова зрялост. Те могат да бъдат доминиращи, рецесивни или свързани с пола. Те обикновено проявяват своя ефект в хомозиготно състояние; в хетерозиготно състояние те намаляват жизнеспособността. Пенетрантността е способността на гена да се проявява фенотипно, изразява се в % и може да бъде пълна или непълна. Пълен – при всички индивиди от една популация, които имат даден ген, той се проявява като признак. Непълен - някои индивиди имат гена, но не се проявяват външно. Експресивността е степента на проява на черта, т.е. една и съща черта се изразява с различна интензивност при различните индивиди.
При наличието на различни летални гени, организмите умират на различни етапи на развитие. По правило леталният ефект на такива гени е рецесивен, т.е. се проявява само когато са в хомозиготно състояние. Когато възникнат мутации с доминиращ летален ефект, организмът умира без да създаде потомство.
Въпреки това има случаи, когато смъртоносни гени, причиняващи видими промени в хетерозиготното състояние, дори стават полезни от икономическа гледна точка. Така сред овцете от породата каракул има животни с красив сребристосив цвят на кожата, който се оценява по-скъпо от обикновения черен каракул.При кръстосването на сиви овце със сиви кочове се оказа, че те винаги са хетерозиготни. При изследване на причините за това явление беше установено, че сред сивите агнета, получени в резултат на такова кръстосване, приблизително 1/3, или около 25% от общото потомство, се разболяха от хроничен тимпанит и умряха. Причината за заболяването са нарушения в дейността на парасимпатиковата нервна система. При кръстосване на сиви кочове с черни овце или сиви овце с черни кочове, потомството дава 50% сиви и 50% черни агнета, а сивите агнета не се разболяват. Оказа се, че в хомозиготно състояние генът, който причинява развитието на сивото оцветяване, има рецесивен летален ефект.
Лисиците имат ценен цвят на козината - платина, причинен от доминантен ген, който в хомозиготно състояние причинява смъртта на ембрионите в ранен стадий на развитие. В такива случаи съотношението на фенотипите във второто поколение се променя, тъй като от трите доминиращи форми умира една (хомозиготна за леталния ген), в резултат на което съотношението на фенотипите става 2:1.
Подобно разцепване е отбелязано при огледалния шаран, при който се наблюдава унаследяване на недоразвитие на люспите: тя се запазва само по средната линия на тялото, останалата част е гола, поради което се нарича линейна. Когато линейните шарани се кръстосват един с друг, потомството винаги се разделя в съотношение 2 линейни: 1 с нормални люспи, тъй като ембрионите, хомозиготни за гена, който причинява недоразвитие на люспите, умират в ранните етапи на развитие.
При пилета, хомозиготни за алела, причиняващ извиване на перата, непълното развитие на перата има няколко фенотипни ефекта. Тези пилета имат недостатъчна топлоизолация и страдат от охлаждане. За да компенсират загубата на топлина, те развиват редица структурни и физиологични адаптации, но тези адаптации са неефективни и смъртността сред такива пилета е висока.
Ефектът на леталния ген се вижда ясно при унаследяването на цвета на козината при мишки. Дивите мишки обикновено имат сива козина, като агути; но някои мишки имат жълта козина. Кръстоските между жълти мишки дават както жълти мишки, така и агути в съотношение 2:1. Единственото възможно обяснение за тези резултати е, че жълтият цвят на козината е доминиращ при агути и че всички жълти мишки са хетерозиготни. Атипичната менделска връзка се обяснява със смъртта на хомозиготни жълти мишки преди раждането. Аутопсията на бременни жълти мишки, кръстосани с жълти мишки, разкрива мъртви жълти малки в техните матки. Ако жълтите мишки и агути бяха кръстосани, тогава нямаше да има мъртви жълти мишки в матките на бременни женски, тъй като при такова кръстосване не може да има потомство, хомозиготно за гена на жълтата вълна.
Промяната във фенотипната сегрегация в съотношение 3:1 във второто поколение на монохибридно кръстосване е свързана с различна жизнеспособност на F2 зиготите. Различната жизнеспособност на зиготите може да се дължи на наличието на летални гени. Летален е ген, който причинява смущения в развитието на организма, което води до неговата смърт или деформация.
Изследването на вродените аномалии показа, че при различни летални гени смъртта на индивидите е различна и може да настъпи на различни етапи от развитието.
Според класификацията, предложена от Rosenbauer (1969), гените, които причиняват смъртта на 100% от индивидите, преди да достигнат зрялост, се наричат летални, повече от 50% се наричат сублетални (полулетални) и по-малко от 50% се наричат субвитални. Все пак трябва да се отбележи, че това разделение е до известна степен произволно и понякога няма ясни граници. Пример за това е свързаната с пола голота при пилетата. Почти половината от голите пилета умират през последните 2-3 дни от инкубацията. От излюпените пилета около половината умират преди 6-седмична възраст, ако се отглеждат при температура от 32-35 ° C. Но ако температурата в брудерите се повиши с 5,5 ° C, тогава значително по-малко голи пилета ще умрат. 4-- След 5 месеца голите пиленца растат с рядко оперение и вече могат да издържат на доста ниски температури.
При естествени условия тази мутация вероятно ще бъде смъртоносна и ще доведе до 100% смъртност на птиците. Горният пример показва, че естеството на проявлението на полулетален ген може до голяма степен да зависи от условията на околната среда.
Смъртоносните гени могат да бъдат доминиращи или рецесивни. Сред първите смъртоносни фактори е открит алелът, който причинява жълтото оцветяване на мишките. Генът на жълтия цвят е доминиращ (Y). Неговият рецесивен алел (y) в хомозиготно състояние причинява появата на черно оцветяване. Кръстосването на жълти мишки една с друга произвежда две части жълти мишки и една част черни, т.е. резултатът е разделяне 2:1, а не 3:1, както следва от правилото на Мендел. Оказа се, че всички възрастни мишки са хетерозиготни (Yy). Когато се кръстосват един с друг, те трябва да са произвели една част от хомозиготното потомство за жълтия цвят (IT), но то умира в ембрионалния период, две части от хетерозиготите (Yy) ще бъдат жълти и една част от хомозиготите за рецесивната черта (yy) ще бъде черна. Схемата за пресичане изглежда така:
По същия начин се наследява сивият цвят на козината при каракулските овце (Соколские, Малич и др.), платиненият цвят при лисиците, разпределението на люспите при линейните шарани и др.
Смъртоносните гени в повечето случаи са рецесивни и следователно могат да останат латентни за дълго време.
Животно, което е напълно здраво и нормално по фенотип, може да бъде носител на летален ген, чийто ефект се открива само при преминаване към хомозиготно състояние. Смъртоносните гени най-често преминават в хомозиготно състояние по време на инбридинг. В практиката на животновъдството при отглеждане на коне има случай на смърт на 25 жребчета на 2-4-ия ден след раждането от ректална деформация - липса на анус (Atresia ani). Оказа се, че всички жребци и кобили, родили такива необичайни жребчета, са произлезли от един и същи жребец. Той беше хетерозиготен за леталния ген (LI). Първоначално този жребец, когато е кръстосан с нормални кобили (LL), ражда потомство, което е нормално по фенотип, но по отношение на генотипа половината от потомството е нормално (LL), а половината са хетерозиготни (LI), носещи рецесивен наклон (0 летален ген. При инбридинг на хетерозиготни животни (Y x Y) се появиха няколко жребчета, хомозиготни за леталния ген (II), с ректална деформация. Всички те умряха.
Известни са десетки аномалии при селскостопанските животни, чиято поява е свързана с рецесивни или доминантни генни мутации. Тези аномалии се срещат в отделните популации с различна честота, която зависи от скоростта на мутационния процес, системата на отглеждане на животните и др. Познаването на специфичните форми на вродени аномалии при животните от всеки вид, както и честотата на тяхното проявление в отделни породи, е необходимо на ветеринарните специалисти за селективна превенция на разпространението на генетична патология.
Аномалии при говедата. Биологичните характеристики на този вид животни са късокосмести и сравнително късно узряване. Една крава обикновено ражда едно теле, което достига полова и физиологична зрялост само до 1,5 години, така че периодът между първото отелване на майката и дъщерята е средно 5 години. В резултат на това появата на необичайно потомство в стадо може значително да намали нивото на възпроизводство и интензивността на селекцията на добитъка. Широка гама от вродена шамалия, определена от летални, полулетални и субвитални гени, е изследвана при говеда. 46 аномалии са включени в Международния списък на леталните дефекти под код А (Таблица 43). Относителната честота на определени видове аномалии във всяка порода или популация може да варира. В Костромската порода, според нашите данни, най-често регистрираната генетична аномалия на главата е скъсяването на челюстта (Таблица 44), в Ярославската порода - синдактилия, в Холмогорската порода - мускулни контрактури, в Черно-и- Бяло - пъпна херния. Аномалиите на централната нервна система са най-често срещаните (21%) при говедата в Германия.
На второ място по честота на регистрация (14%) е сложна аномалия - комбинация от пъпна херния с цепка на корема и плода като цяло. Честотата на аномалиите или процентът на анормалното потомство към общия брой в рамките на определени популации също може да бъде много различна и според средните оценки не надвишава 1%. Този индикатор обаче зависи от пълнотата и точността на записване на аномалии. Така в Германия, след организиране на ясно счетоводство, заключиха, че честотата на аномалиите се е увеличила няколко пъти. Въпросът е: всички аномалии подлежат ли на визуално наблюдение? Очевидно не всички. Така в костромската порода средната честота на всички форми на аномалии за 12-годишен период е 1,15%. Честотата на общата смъртност на котилото (абортирани, мъртвородени, необичайни, мъртви телета без видими дефекти) в тази ферма е 10,2%. Определена част от тази смъртност е свързана и с генни мутации, които причиняват не морфологични дефекти, а метаболитни нарушения и други аномалии, идентифицирането на които е възможно само чрез специални методи.
Производителите могат да играят специална роля в разпространението на генетични аномалии както при говеда, така и при други животински видове. От всеки баща с изкуствено осеменяване могат да се получат стотици и хиляди приплоди годишно. Така от един бик в чужбина са получени 100 хиляди телета. Ако такъв баща се окаже носител на генна мутация, тя бързо ще се разпространи в породата. Ето няколко примера от множеството факти, описани в литературата. В резултат на интензивното използване на бика Принц Адолф, докаран в Швеция, и последвалото спонтанно инбридинг върху него, честотата на обезкосмяване в някои шведски стада е над 5%. Същата ситуация възникна и в Швеция след вноса на бика Gallus, който се оказа хетерозиготен носител на гена, причиняващ липсата на крайници.
В потомството на отделни бикове от черно-шарени и шароле в САЩ и Германия са регистрирани случаи на раждане на телета джуджета с честота съответно 23,3 и 22,2%. В първия В Чехословакия при изследване на потомците на 166 бащи е установено, че 43 от тях са носители на смъртоносни гени. При един бик - носител на доминиращата аномалия "цепнатина на устната" - дефектът се проявява сред 44% от биковете и 71% от юниците от неговото потомство.
В костромската порода анализирахме разпространението на скъсяването на долната челюст и мопсовия вид чрез бика Бурхан, който самият имаше дефектни телета в потомството си; неговите синове, внуци, правнуци, потомци по женски също са дали ненормално потомство (фиг. 57). Повечето телета са получени от инбридинг и чифтосване на родители с нормален фенотип и общ прародител. Следователно можем да заключим, че тази аномалия има рецесивен начин на наследяване. Фигурата показва, че най-голям брой дефектни телета са регистрирани в поколението на бика Zheton 3501 (внук на бика Burkhan) при използване в търговска ферма, където някои от кравите имат същия рецесивен ген в генотипа си.
Аномалии при свинете. Международният списък на смъртоносните дефекти при свинете включва 18 генетични аномалии. Основната част от тях се причиняват от автозомно рецесивни гени (Таблица 45). Генетичните аномалии могат да играят значителна роля в патологията на свинете. Нека да разгледаме няколко такива примера. В Испания изследване на 23 449 прасенца от 2 399 котила, получени от нерези от породите Duroc, Yorkshire, Hampshire и White Chester, разкрива съответно 6,21; 6,02; 9,66; 2^62% от ненормалните котила.
Според Оливие (1979) 7 генетични аномалии на кожата, 17 на скелета, 3 на очите, 13 на нервно-мускулната, 6 на кръвта, 6 на хормоналния метаболизъм, 5 на храносмилателната система и 9 на пикочно-половата система са описани при прасета. Основните аномалии са крипторхизъм, херния, псевдохермафродитизъм и др. Авторът на изследването смята, че тези аномалии са резултат от действието на един ген на различни етапи от формирането на ембриона.
В Дания 6669 мъртви прасенца от 2936 котила са изследвани за период от две години, за да се определи естеството и честотата на вродените аномалии. Различни аномалии са открити при 1,4% от родените прасенца или 6,2% от убитите преди отбиването. По време на следкланичен преглед, 25,9% от анормалните прасенца са с недоразвити клапи, незатворен анус, субаортна стеноза, ектопия на сърцето и други дефекти на сърдечно-съдовата система. При 23,4% от прасенцата са открити различни нарушения в развитието на двигателната система. Аномалии на централната нервна система са открити при 5,9% от прасенцата, включително раздвоен мозък и мозъчно хидроцеле. Свръхрастеж на ректума, тънките черва или тяхното непълно развитие са установени при 30% от прасенцата, а различни хернии и асцит - при 6,8%. „Цепка на устната“, цепка на небцето, риноцефалит и други аномалии на лицевата част на главата са установени при 6,1%; хермафродитизъм, цепнатина на уретера, хидроцеле на бъбреците и уретрата - при 1,7% от прасенцата. Тези аномалии са се появили в потомството на отделни бащи по време на инбридинг, което показва наследствения характер на тяхното възникване.
Много убедителни доказателства за наследствения характер на крипторхизма при прасетата са получени от Фридин и Нюман. Според техните данни в Канада едностранен и двустранен крипторхизъм се наблюдава годишно при 1-2% от всички глигани, които влизат на пазара. Авторите са кръстосали крипторхидите с техните майки и пълни сестри. Потомството от такива кръстоски се чифтосва помежду си. В резултат на такава селекция и селекция, честотата на крипторхизъм при експериментални животни от породата йоркшир се увеличи средно до 42,9% и особено когато се използват двама производители. При изследване на прасенца в Съединените щати за една година бяха открити около 400 хиляди животни със скротална херния.
Наблюденията показват, че причината за нарушената плодовитост при нерезите често е хипоплазия на тестисите. Честотата на тази аномалия, според изследователи от Германия, е 19,6%. За размножаване са оставени 30 такива нереза
Va, всеки от тях е покривал от 4 до 40 кралици (общо 439 глави), но само четири от тях са родили потомство. Анализът показа, че патологичните форми на спермата при тези нерези са 80-100%. Всичките 30 анормални животни са имали общи предци, което показва наследствения характер на хипоплазията на тестисите и дефектите на спермиогенезата.
Наличието на кратерни зърна при прасетата е един от сериозните дефекти, тъй като прасенцата не получават мляко от тях. Според Баварския институт по животновъдство (Германия) честотата на тази аномалия при немските ландраси е 6,6%. Както отбелязват П. Н. Кудрявцев и др. (MVA), броят на прасетата с неактивни кратерни биберони се е увеличил през последните години. Броят на тези зърна варира от 1 до 8. Прасенцата, които получават кратерни зърна, умират.
Кратерността е черта, причинена от един автозомно рецесивен ген. Това е проверено експериментално от П. Н. Кудрявцев и др. След като преди това са идентифицирали нерези и женски свине, които са носители на аномалии (KchKch), нормални, но хетерозиготни индивиди (KchKch), които произвеждат прасенца с кратер в потомството си, и нормални хомозиготни прасета (KhKch), авторите извършват кръстосване между тези групи животни . В първия вариант са кръстосани 27 нормални хомозиготни майки с 15 глигана. Всички 258 потомци бяха нормални. Във втория вариант, където единият от родителите е хомозиготен (KchKch), а другият е хетерозиготен (Kchkch), всички прасенца също са нормални. В третия вариант са кръстосани 13 хетерозиготни нереза с 16 хетерозиготни майки. От 168 родени прасенца, 39 (23,2%) са имали кратерни зърна. И накрая, в четвъртия вариант, единият от родителите е хомозиготен, а другият е хетерозиготен. Те произведоха 170 прасенца, от които 86 (50,5%) бяха с нормални зърна и 84 (49,5%) с кратерни зърна. Резултатите от този експеримент доказват рецесивния начин на унаследяване на кратера на зърното при прасетата.
Аномалии при овцете. При овцете са описани около 90 вродени аномалии. Според Dennis и Leipold, повечето известни генетични дефекти при овцете са причинени от моногенен автозомно-рецесивен начин на унаследяване (Таблица 46). Най-често срещаните краниофациални дефекти при този вид животни са краниофациални дефекти, особено агнатия, както и изкривяване на предните крайници, микроагнатия, хермафродитизъм, крипторхизъм, хипоспадия, прогнатия, анална атрезия, микротия, ентропия, тортиколис, полителия, артрогрипоза. Анализът показа, че 55,4% от дефектите са свързани с опорно-двигателния апарат, 12,7 - с храносмилателната система, 9,7 - със сърдечно-съдовата система, 7,1 - с урогениталната система, 6 - с централната нервна система, 3,5 - с аномалия. на връзките, 3,2 - към коремната, 1,5% - към ендокринната система. Въпреки че честотата на отделните дефекти е ниска, кумулативният принос на всички аномалии може да причини щети на фермите. В Нова Зеландия, страна с развито овцевъдство, честотата на смъртоносни дефекти е около 1% от мъртвите агнета. В САЩ летални дефекти през първите 2 седмици от живота са наблюдавани при 11,4% от агнетата.
Средната ембрионална смъртност при овцете е 20%. Това показва, че много неидентифицирани летални гени може да са активни през този период.
Индивидуалните генетични аномалии при овцете могат да бъдат широко разпространени. Така в България при стада от мериносови овце се наблюдава висока смъртност на агнетата в ранния постнатален период. Това се дължи на факта, че агнетата не получават мляко поради дефекти на вимето на майките си: от хипоплазия с малки остатъци от жлезиста тъкан до пълното й отсъствие. Честотата на тази наследствена аномалия в различни стада варира от 6 до 40%.
При овцете често се наблюдава крипторхизъм, който е съчетан с такава вторична сексуална характеристика като полюс. Опилените кочове имат ниска плодовитост. Чрез строга селекция за разплод беше възможно да се създаде тип овни с нормална плодовитост, но сред тях се срещат и необичайни индивиди.
Аномалии при птиците. Птиците, предимно пилетата, са най-изследвани във връзка с генетиката на аномалиите. Международният списък на леталните дефекти включва 45 аномалии при пилетата, 6 при пуйките и 3 при патиците. Най-честите аномалии на клюна са (клюн на папагал, кръстосан клюн). Честотата им, според William et al., е 1,1% от загубите по време на инкубация на яйца от пилета White Leghorn и Rhode Island. Аномалиите на клюна също са често срещани при патиците.
Макс Гибън и Шакелфърд описаха аномалия при кръстосване на бели легхорни с бутеркамп и бантам и последващо размножаване „сами по себе си“ - полидактилия. В допълнение, синдактилия и оперени крака са наблюдавани при пилета. Честотата на синдрома е 16,8%. При кръстосването на анормален F2 петел с фенотипно нормални кокошки се наблюдава разделяне - половината нормални и половината анормални индивиди (1:1). Установено е, че този комплекс от признаци се контролира от един автозомен ген, който има полулетален ефект, тъй като ембрионалната и постембрионалната преживяемост на анормалните пилета е много ниска.
Аномалии при конете. От наследствените аномалии при конете 10 са включени в Международния списък на смъртоносните дефекти. Сред тях са 3 аномалии на скелета, 2 на репродуктивната система, 2 на бъбреците и мускулите, по една аномалия на червата, нервната система и органите на зрението.
При коне от впрегатни породи атрезия на дебелото черво е по-честа. Разпространението на тази аномалия е отбелязано в потомството на жребеца Superba от породата Percheron. Описано е и при жребчета от чистокръвна ездитна порода от източноафрикански произход. При впрегатните породи е регистрирана несъвършена епителиогенеза. Атаксия, наречена Олденбург, е открита в жребчета от Олденбург в Германия. Разпространява се в ред 9. Една от най-често регистрираните аномалии при конете е пъпната херния. Среща се при леки и тежки породи.
При конете са известни няколко други генетични и наследствено-средови аномалии. Така в САЩ коне от няколко породи имат жребчета със специфично бяло петно, наречено „оверо“. При кръстосване на коне от типа "overo" се раждат жребчета с розова кожа, при които се наблюдават хипоплазия на чревния тракт и изоеритролиза, както и колики, водещи до смърт.
В Англия са регистрирани и изследвани животни с нарушена координация на движенията - "болест на колебание" при коне от полукръвната порода за езда. Установена е генетична предразположеност към тази аномалия.
Наследствеността на дерматозите на крайниците при конете е доказана. Дефекти с наследствена предразположеност са често срещаните хронични деформиращи възпаления на скакателната става - лопатата, т. нар. „крак на патерицата” при жребчетата, както и хроничното асептично възпаление на венечния блок на копитата, наблюдавано предимно при състезателни и състезателни коне. коне.









- Движимо имущество, придобито след 01
- Данък върху имуществото, придобито след 01
- Морско конче - доклад за съобщения Откога съществува морското конче на земята
- Ванилови кексчета с млечен шоколад
- Рецепта за палачинки с кисела мая със снимка
- История на изобретяването на водка
- Наследството на светите отци: православни притчи за живота и морала и библейски цитати
- Кралско месо, новогодишна рецепта за готвене на кралско месо Кралско месо с картофи във фурната
- Бял квас за окрошка Направете квас за окрошка у дома
- Химия (Атомен строеж) (презентация)
- Традиции и обичаи на Англия, които ще ви изненадат
- Наследственост и развитие Толкова ли е важна наследствеността?
- Книги за Хари Потър на английски
- Организационни фактори, предизвикващи стрес
- Рецепта за задушено зеле в бавна готварска печка: проста, бърза и вкусна
- Осмица жезли, описание и характеристики на картите
- Стек и медальони от сьомга
- Обобщение на индивидуален логопедичен урок за звукова автоматизация Обобщение на звукова автоматизация l в изречения
- Нижегородски държавен педагогически университет на име
- Всички, които разпънаха Христос, умряха от ужасна смърт Пилат Понтийски наистина ли беше там?