Genes recesivos, dominantes, letales, mutaciones. III.2.8
Hasta ahora hemos considerado la genética humana por tipos de herencia; ahora pasaremos a una categoría muy importante de genes, a saber, los genes letales y subletales. Los genes letales y subletales son una clase de mutaciones con efectos negativos marcadamente pronunciados sobre el desarrollo. Como veremos más adelante, en muchos casos las mutaciones agravan y destruyen la herencia de una persona. Este


el agravamiento por mutaciones es especialmente pronunciado bajo la influencia de genes letales.
Los alelos letales y subletales pueden ser dominantes o recesivos. En el caso de letalidad dominante, el niño muere en el período prenatal o en la primera infancia. Está bastante claro que esos genes no se pueden heredar. Su presencia indica la aparición de mutaciones. La frecuencia total de tales letales dominantes no es tan pequeña. Su número es especialmente grande bajo la influencia de la radiación. En la genética de ratones y Drosophila, los letales dominantes se utilizan ampliamente al analizar la dependencia de la frecuencia de mutación de la dosis y al analizar otras características de los efectos mutagénicos. En presencia de genes subletales, también llamados genes semiletales, el niño muere en la primera infancia o algo más tarde, pero, por regla general, antes de llegar a la pubertad. Un ejemplo de mutación subletal dominante es la epiloia, una enfermedad caracterizada por el efecto de todo un síndrome de síntomas negativos. Los niños con epiloia tienen crecimientos patológicos de la piel, se caracterizan por retraso mental, epilepsia, tumores del corazón, riñones y otros órganos. En casos raros, los pacientes con epiloia sobreviven y se casan con un cónyuge sano. La mitad de los hijos de un matrimonio de este tipo padecen epiloia congénita (Fig. 131).
Otro ejemplo sorprendente de una mutación dominante subletal es la enfermedad del retinoblastoma. Este


el gen dominante provoca un tumor canceroso en el ojo, que casi siempre provoca la muerte del niño a una edad temprana.
Sin embargo, la mayor parte de las deformidades congénitas recae en letales y subletales recesivas. En este caso, los genes dañinos recesivos están ocultos en el genotipo de padres aparentemente sanos; sin embargo, cuando dos de estos heterocigotos se casan, aproximadamente el 25% de sus hijos resultan estar enfermos.
La Figura 132 muestra los principales tipos de cruces en poblaciones humanas que se realizan al heredar genes letales y subletales recesivos. al cuadrado 1 Presenta el tipo de matrimonios entre personas genéticamente normales. Tanto en los padres como en todos sus descendientes, todos los alelos están representados por genes normales (alelo +). al cuadrado 2 Representa un matrimonio entre una persona genéticamente sana y un heterocigoto (símbolo a). En este caso, el alelo negativo se transmite a la mitad de la descendencia, aunque permanece oculto. al cuadrado 3 una persona genéticamente sana se casa con una persona enferma (símbolo Automóvil club británico), Todos los niños son aparentemente sanos, pero en el estado heterocigoto portan una mutación subletal. A. al cuadrado 4 un heterocigoto se casa con un paciente; La mitad de sus hijos están aparentemente sanos pero portan una mutación subletal, la otra mitad está enferma. Finalmente, un cuadrado 5 en caso de matrimonio de dos personas enfermas (símbolos Automóvil club británico) toda la descendencia estará enferma. Las enfermedades congénitas causadas por la acción de genes recesivos letales y subletales incluyen la idiotez amaurótica infantil, en la que el tejido cerebral degenera, se produce ceguera y el niño muere; patología congénita de la piel con la presencia
grietas sangrantes profundas - ictiosis (Fig. 133); parálisis infantil congénita y una serie de otras enfermedades congénitas.
Muchos abortos espontáneos tempranos son causados por la fuga de genotipos letales.
Se conocen mutaciones dominantes que en los heterocigotos determinan algún rasgo más o menos neutro, que, sin embargo, tienen un efecto letal en el estado homocigoto. Por ejemplo, el alelo de pigmentación del xeroderma en estado heterocigoto provoca pecas graves. Sin embargo, los homocigotos desarrollan xeroderma pigmentoso. Se trata de una enfermedad subletal (Fig. 134), en la que la acción de la luz provoca daños en la piel, que luego conducen a un crecimiento maligno.
Uno de los alelos en una persona en estado heterocigoto provoca un exceso de colesterol en la sangre. Sin embargo, los niños homocigotos para este alelo desarrollan una anomalía subletal grave.
En todos estos casos, tenemos un cuadro clásico de herencia de dominantes con efecto letal recesivo, previamente establecido en experimentos con Drosophila, ratones y otros organismos. Por ejemplo, el ratón tiene un alelo bien conocido. Y, lo que hace que la piel se vuelva amarilla en los heterocigotos. Sin embargo, todos los intentos de obtener una raza pura de ratones amarillos fracasaron. Al cruzar dos ratones amarillos, en promedio, dos ratones amarillos siempre producen uno de diferente color, por ejemplo, negro. La naturaleza de este fenómeno se explica por el hecho de que el gen del color amarillo dominante resultó ser
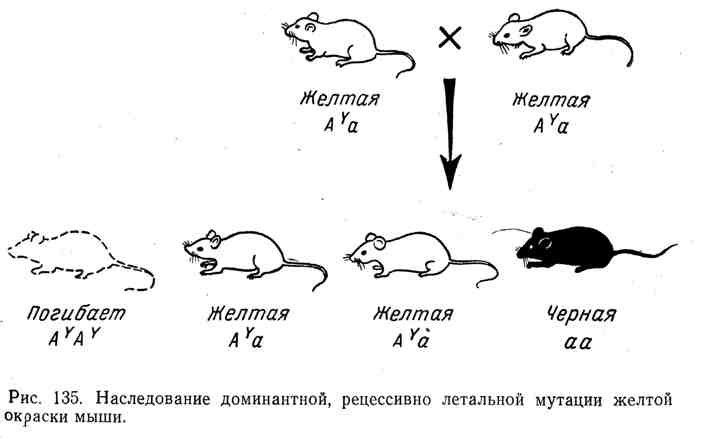
letal recesivo. Todos los homocigotos( AY AY) fallecido. El ratón amarillo siempre es heterocigoto, junto con el alelo.Y contiene otro alelo, p. A(de color negro). El curso de la herencia al cruzar heterocigotos. A y a y dividir según la fórmula 2: 1 se muestran en la Figura 135.
- Fuente-
Dubinin, N.P. Horizontes de la genética / N.P. Dubinin. – M.: Educación, 1970.- 560 p.
Vistas de publicaciones: 121
La viabilidad desigual de cigotos de diferentes genotipos puede estar asociada con mutaciones letales dominantes o recesivas que surgen en los gametos de los individuos parentales. Pueden realizarse en diferentes etapas de la embriogénesis o en el período postembrionario. Naturalmente, la muerte de algunos cigotos afecta la naturaleza de la escisión.
Influencia en la división de genes dominantes con efectos letales recesivos.
Los genes dominantes con efectos letales recesivos tienen un efecto pleiotrópico; por un lado, determinan el carácter dominante de la manifestación de cualquier rasgo en un heterocigoto, por otro, provocan la muerte de individuos en un homocigoto, es decir, manifiestan su efecto letal como recesivo. Estos genes se conocen en muchos animales: coloración amarilla en ratones, patas cortas en pollos, escala lineal en carpas, coloración platino en zorros, coloración shirazi en ovejas y muchos otros. Provocan una desviación de la división 3:1, convirtiéndola en una división 2:1. El análisis de segregaciones que involucran genes dominantes con efectos letales recesivos se complica por el hecho de que en muestras relativamente pequeñas, no siempre es posible distinguir entre divisiones 3:1 y 2:1, y el método estadístico de prueba utilizando el criterio χ 2 no permite elegir entre hipótesis. Por ejemplo, si al cruzar ratones amarillos entre sí se obtiene una división de 65 amarillos: 32 negros, el método %2 no rechaza dos hipótesis: 1 - la división en el experimento corresponde a una división de 3:1, χ2 = 3,29, p>0,05; 2 - la división en el experimento corresponde a la división 2:1, χ 2 = 0,17, p>0,05.
Sólo en muestras grandes se distingue una división 2:1 de una división 3:1.. Al resumir los datos sobre la herencia del color amarillo en ratones obtenidos por diferentes investigadores, la división al cruzar ratones amarillos entre sí fue 2386 amarillos: 1235 negros - ∑ = 3621, χ 2 = 0,96 en H 0 - 2:1 ( en H 0 - 3:1 en este caso χ 2 = 160!).
Es obvio que diferentes métodos de pruebas genéticas, por ejemplo, la realización de análisis y retrocruzamientos, pueden contribuir a la solución correcta del problema de la herencia. En el ejemplo analizado anteriormente, estas son cruces: amarillo de F 1 × negro; amarillo de F 1 × amarillo P. En el primer caso la división debe ser 1:1, en el segundo nuevamente 2:1.
Es más difícil descubrir que existe una desviación y que se debe a una viabilidad diferente de los cigotos cuando la muerte de algunos cigotos ocurre en la etapa postembrionaria. En este sentido, es necesario monitorear la fertilidad y mortalidad de la descendencia si existe un supuesto de letalidad. La base para el supuesto de letalidad es la imposibilidad de obtener descendencia uniforme al criar individuos de un determinado fenotipo. En los problemas III.2 y III.3 se dan ejemplos de análisis.
Problema nº III.2
En la descendencia de cruces de visones de marta plateada entre sí en F 1, siempre se observa división: aparecen individuos de marta plateada y marrón.
En un experimento, varias camadas de cruces de marta plateada produjeron 93 cachorros de marta plateada y 43 marrones, con un tamaño promedio de camada de 3,66 cachorros. Al cruzar el sable plateado con visones marrones, se obtuvieron 39 martas marrones y 41 sables plateados. Al cruzar visones marrones entre sí, solo se obtuvo descendencia marrón I. El tamaño de la camada en los dos últimos cruces fue de 4,9 a 5,2 cachorros.
Explique la división, determine los genotipos de los visones marta plateada y marrón.
Análisis
Dado que los visones de marta plateada siempre producen segregación cuando se cruzan entre sí y con los marrones, obviamente son heterocigotos. Para determinar la cantidad de genes, considere las divisiones en el experimento:
H 0 - diferencias en un gen, división 3:1, χ 2 = 3,2, p>0,05. La desviación es aleatoria, la hipótesis no se rechaza.

La relación corresponde bien a una división 1:1 para un gen, χ 2 = 0,05, p>0,80. La desviación es aleatoria, la hipótesis no se rechaza.
Una comparación de los datos sobre la fertilidad de los visones en diferentes cruces indica una muerte parcial de los cigotos al cruzar visones de marta plateada. Podemos suponer la muerte de los homocigotos dominantes. Entonces el genotipo de los visones de marta plateada es Aa, los marrones - aa, y la división en la cruz no es 3:1, sino 2:1 (χ 2 = 0,17, p>0,05). Para la confirmación final de esta hipótesis, es necesario realizar un cruce adicional entre individuos de marta plateada para aumentar la muestra y probar estadísticamente la hipótesis 2:1. En algunos casos es necesario realizar análisis más complejos.
Problema nº III.3
En cruces de zorros platino, de cara blanca y blanco georgiano con zorros negros plateados, se encontró que los colores platino, de cara blanca y blanco georgiano, que provocan un debilitamiento general de la pigmentación y la aparición de varios píos, no son sexo. -vinculado. El cruce de cada uno de estos mutantes con negros plateados dio como resultado una segregación 2:1, siendo el color negro plateado recesivo. En consecuencia, cada uno de ellos está controlado por un gen dominante con un efecto letal recesivo. La muerte de algunos mutantes se evidencia en los datos sobre el tamaño de la camada: los negros plateados tienen 4,5 cachorros por camada, los de cara blanca 3,5, los platino y los blancos georgianos, menos de 3,5.
Para establecer si estas mutaciones afectaban a un gen o a diferentes, se realizaron cruces cuyos resultados se detallan a continuación. La prueba de alelismo no es aplicable en este caso, ya que las mutaciones son dominantes:

Explique la división, determine el genotipo de todas las formas.
Análisis
1. Las divisiones en los cruces 1 y 2 corresponden a la relación 1:1:1:1 (χ 2 = 5,83, p>0,10 para el cruce 1, χ 2 = 0,55, p>0,90 para el cruce 2). Pueden ser el resultado de varias razones.
1. Dado que en los cruces 1 y 2 existen 4 clases de escisión y una proporción de 1:1:1:1, se puede suponer que las formas estudiadas se diferencian en dos genes dominantes heredados independientemente con un efecto letal recesivo que interactúan según el tipo de complementariedad. En este caso, uno de los genes está representado por dos alelos dominantes diferentes.

2. La segregación 1:1:1:1 puede ser consecuencia del estrecho vínculo de estos dos genes en ausencia de cruce entre ellos:

(En las redes de Punnett, se dan radicales fenotípicos, genes que manifiestan su efecto en el fenotipo).
3. Las diferencias de color son causadas por tres genes dominantes heredados independientemente con un efecto letal recesivo. En los cruces primero (a) y segundo (b), la división ocurre en dos genes diferentes.

4. La segregación 1:1:1:1 puede resultar del estrecho vínculo de tres genes que interactúan en ausencia de entrecruzamiento.

* (El orden de los genes puede ser diferente; se da de forma arbitraria.)
5. La segregación en la proporción 1:1:1:1 puede ser el resultado de diferencias en un gen, representado por una serie de cuatro alelos, tres de los cuales son dominantes con un efecto letal recesivo y el cuarto es recesivo:

Para elegir entre estas hipótesis, se hizo una cruz: blancos con negros plateados.
Si un rasgo estuviera controlado por dos o tres genes, entonces con su herencia independiente se esperaría la aparición de cuatro clases fenotípicas:

Se debería obtener un resultado similar al cruzar blancos obtenidos de zorros blancos platino o georgianos con negros plateados, sin embargo, en lugar de cara blanca deberían haber aparecido los platino, lo que se puede comprobar fácilmente escribiendo los cruces correspondientes.
La división obtenida en el cruce de prueba (la aparición de los zorros de cara blanca y los zorros blancos georgianos) puede explicarse por la interacción de dos (o tres) genes estrechamente relacionados o por la interacción de tres alelos con una diferencia monogénica entre los originales. formas.

La fuerte disminución de la viabilidad de los zorros blancos observada en el experimento habla a favor de la acción de las mutaciones alélicas, ya que en este caso los zorros blancos son genotipos compuestos de dos mutaciones dominantes de un gen, ambas con un efecto letal recesivo. Es difícil esperar una disminución de la viabilidad cuando interactúan dos genes diferentes (complementariedad). Por lo tanto, concluyeron que los colores blanco, cara blanca, blanco georgiano, platino y negro plateado en los zorros están controlados por una serie de alelos de un gen, tres de los cuales son dominantes con un efecto letal recesivo. Genotipos de zorro: blanco A 1 / A 2, A 1 / A 3, A 2 / A 3; cara blanca A 1/a; platino A 3/a; Blanco georgiano A 2/a; negro plateado a/a (según Belyaev et al., 1973).
Cabe recalcar que con alelismo múltiple en el caso de heterocigosidad de las formas originales, el número máximo de clases fenotípicas en la división puede no ser 3, sino 4, como en el caso descrito anteriormente. En una población, el número de posibles genotipos con alelismo múltiple aumenta muchas veces; se puede determinar mediante la fórmula: 1 / 2n (n+1), donde n es el número de alelos. Por ejemplo, si hay 7 alelos para un locus, el número de genotipos posibles en la población será 28: 1/2 × 7 × 8 = 28.
Efecto de las mutaciones letales recesivas sobre la segregación.
Moscas autosómicas recesivas y ligadas al sexo, Al provocar la muerte de homocigotos en cruces de heterocigotos letales, pueden influir en la división de genes vinculados a letales. En este caso, la proporción de descendientes en la división depende de la distancia entre el gen en estudio y el letal, así como de la distancia. Según el tipo de heterocigoto, en los genes de posición cis o trans se introducen en los heterocigotos. Para identificar letales se suelen realizar varios cruces de prueba. Un ejemplo de análisis de vuelo es el problema No. III.4.
Problema nº III.4
En la línea Drosophila de la línea No. 100, que contenía inversiones, la mitad de las hembras tenían un color de cuerpo gris, la otra mitad amarillo, y las hembras amarillas resultaron ser estériles. Todos los machos de esta línea eran de color amarillo. La proporción entre mujeres y hombres era diferente de lo normal, más bien una división de 2♀♀:1♂♂. Se ha sugerido que la falta de machos en la línea número 100 probablemente se deba a la presencia de hembras grises en el cromosoma X, que son obviamente heterocigotas: la línea se divide en hembras grises y amarillas. Para establecer la estructura genética de esta línea y comprobar la suposición de la presencia de letal en el cromosoma X de las hembras, se realizaron cruces, cuyos resultados se presentan a continuación.
Cruces recíprocas

hembras grises de la F 1 fueron individualmente cruzado con machos amarillos de la línea No. 100.

Análisis
Según el análisis, anotaremos los esquemas de todos los cruces.

Las hipótesis propuestas explican bastante bien todos los resultados. Sin embargo, la cuestión del motivo de la esterilidad de las hembras amarillas de la línea N° 100 sigue sin estar clara. Invitamos al lector a reflexionar sobre esta cuestión y proponer algunas hipótesis para explicarla.
Ud. plantas Muchas mutaciones recesivas están asociadas con la falta o ausencia de clorofila, lo que conduce a una disminución de la viabilidad de las plantas o a su muerte en diversas etapas de desarrollo. Esto causa desviaciones en la división y también hace necesario tener en cuenta la división no solo en las plántulas, sino también en etapas posteriores de desarrollo para determinar la proporción de muerte de la planta y la naturaleza de la herencia del rasgo. Por tanto, el maíz es homocigoto para el gen. wd (deficiencia de blanco) tener plántulas blancas (divididas en plántulas 3/4 verdes: 1/4 blancas). Sin embargo, después de 1 a 3 semanas, todas las plantas blancas mueren después de agotar las reservas de nutrientes de la semilla y en las últimas etapas del desarrollo de la planta, la división desaparece: 3 verdes: 0 blancas. Se conocen mutaciones similares en guisantes, cebada, centeno, trigo, etc.
Otras mutaciones provocan la muerte sólo de una parte de los individuos en una determinada etapa de desarrollo, lo que conduce a una disminución de la proporción de recesivos en la división y a un cambio en la proporción de fenotipos: 4:1, 5:1, etc. Estas proporciones varían, ya que, por regla general, la viabilidad de tales mutantes depende en gran medida de las condiciones.
Inhumanos La reducción de la viabilidad y la letalidad debido a la acción de mutaciones recesivas se manifiesta en diferentes períodos de embriogénesis y en diferentes etapas de desarrollo. Las razones de la disminución de la viabilidad y el efecto letal pueden estar asociadas tanto con mutaciones genéticas como con anomalías cromosómicas. El análisis citogenético de embriones abortados nos permite determinar la causa de la muerte de muchos de ellos. En promedio, debido a aberraciones cromosómicas en todas las etapas del embarazo, se producen más del 42% de los abortos espontáneos: una proporción significativa de los recién nacidos con aberraciones cromosómicas muere durante el primer año de vida y los siguientes.
Entre las mutaciones genéticas letales que provocan la muerte fetal o en la infancia, se pueden nombrar mutaciones recesivas que provocan talasemia, anemia falciforme, fibrosis quística, ictiosis congénita, anencefalia (ausencia de cerebro), fenilcetonuria, etc.
Para estudiar mutaciones letales o reductoras de la viabilidad en humanos, se utilizan ampliamente métodos de análisis citogenético y bioquímico, el estudio de la estructura y actividad de las enzimas en la salud y la enfermedad, así como en portadores heterocigotos; cromatografía, diferentes tipos de electroforesis.
Los genes letales son genes mutacionales que provocan la muerte de un individuo antes de que alcance la madurez sexual. Pueden ser dominantes, recesivos o ligados al sexo. Generalmente exhiben sus efectos en el estado homocigoto; en el estado heterocigoto reducen la viabilidad. La penetrancia es la capacidad que tiene un gen de manifestarse fenotípicamente, se expresa en % y puede ser completa o incompleta. Completo: en todos los individuos de una población que tienen un gen determinado, este se manifiesta como un rasgo. Incompleto: algunos individuos tienen el gen, pero no se manifiestan exteriormente. La expresividad es el grado de manifestación de un rasgo, es decir el mismo rasgo se expresa con diferente intensidad en diferentes individuos.
En presencia de diferentes genes letales, los organismos mueren en diferentes etapas de desarrollo. Como regla general, el efecto letal de tales genes es recesivo, es decir. se manifiesta sólo cuando están en un estado homocigoto. Cuando se producen mutaciones con un efecto letal dominante, el organismo muere sin producir descendencia.
Sin embargo, ha habido casos en los que genes letales, que provocan cambios visibles en el estado heterocigoto, incluso resultan útiles desde el punto de vista económico. Así, entre las ovejas de raza Karakul hay animales con un hermoso color de piel gris plateado, que se valora más que la raza Karakul negra habitual. Al cruzar ovejas grises con carneros grises, resultó que siempre son heterocigotos. Al estudiar las causas de este fenómeno, se encontró que entre los corderos grises obtenidos como resultado de dicho cruce, aproximadamente 1/3, o alrededor del 25% de la descendencia total, enfermó de timpanitis crónica y murió. La causa de la enfermedad fueron alteraciones en la actividad del sistema nervioso parasimpático. Cuando se cruzaron carneros grises con ovejas negras o ovejas grises con carneros negros, la descendencia produjo un 50% de corderos grises y un 50% de negros, y los corderos grises no enfermaron. Resultó que en el estado homocigoto el gen que provocaba el desarrollo de la coloración gris tenía un efecto letal recesivo.
Los zorros tienen un valioso color de pelaje: el platino, causado por un gen dominante que, en estado homocigótico, provoca la muerte de los embriones en una etapa temprana de desarrollo. En tales casos, la proporción de fenotipos en la segunda generación cambia, ya que de las tres formas dominantes, una (homocigota para el gen letal) muere, como resultado de lo cual la proporción de fenotipos se vuelve 2:1.
Una división similar se observó en la carpa espejo, en la que se observó herencia de subdesarrollo de escamas: se conservó solo en la línea media del cuerpo, el resto estaba desnudo, por eso se le llama lineal. Cuando se cruzan carpas lineales entre sí, la descendencia siempre se divide en una proporción de 2 lineales: 1 con escamas normales, ya que los embriones homocigotos para el gen que causa el subdesarrollo de las escamas mueren en las primeras etapas de desarrollo.
En pollos homocigotos para el alelo que causa el rizado de las plumas, el desarrollo incompleto de las plumas tiene varios efectos fenotípicos. Estos pollos tienen un aislamiento térmico insuficiente y sufren frío. Para compensar la pérdida de calor, desarrollan una serie de adaptaciones estructurales y fisiológicas, pero estas adaptaciones son ineficaces y la mortalidad es alta entre estos pollos.
El efecto de un gen letal se ve claramente en la herencia del color del pelaje en ratones. Los ratones salvajes suelen tener pelaje gris, como el agutí; pero algunos ratones tienen pelaje amarillo. Los cruces entre ratones amarillos producen tanto ratones amarillos como agutí en una proporción de 2:1. La única explicación posible para estos resultados es que el color del pelaje amarillo es dominante en los agutíes y que todos los ratones amarillos son heterocigotos. La relación mendeliana atípica se explica por la muerte de ratones amarillos homocigotos antes de nacer. Las necropsias de ratones amarillos preñados cruzados con ratones amarillos revelaron crías amarillas muertas en sus úteros. Si se cruzaran ratones amarillos y agutíes, entonces no habría ratones amarillos muertos en el útero de las hembras preñadas, ya que con tal cruce no puede haber descendencia homocigótica para el gen de la lana amarilla.
Un cambio en la segregación fenotípica en una proporción de 3:1 en la segunda generación de un cruce monohíbrido se asocia con una viabilidad diferente de los cigotos F2. La diferente viabilidad de los cigotos puede deberse a la presencia de genes letales. Letal es un gen que causa alteraciones en el desarrollo de un organismo, lo que conduce a su muerte o deformidad.
El estudio de las anomalías congénitas ha demostrado que con diferentes genes letales, la muerte de los individuos es diferente y puede ocurrir en diferentes etapas de desarrollo.
Según la clasificación propuesta por Rosenbauer (1969), los genes que provocan la muerte del 100% de los individuos antes de que alcancen la madurez se denominan letales, más del 50% se denominan subletales (semi-letales) y menos del 50% se denominan subvitales. Sin embargo, cabe señalar que esta división es hasta cierto punto arbitraria y, en ocasiones, no tiene límites claros. Un ejemplo es la desnudez ligada al sexo en las gallinas. Casi la mitad de los polluelos desnudos mueren en los últimos 2 o 3 días de incubación. De los polluelos nacidos, aproximadamente la mitad muere antes de las 6 semanas de edad si se crían a una temperatura de 32-35 °C. Pero si la temperatura en las criadoras aumenta en 5,5 °C, morirán muchos menos polluelos desnudos. 4-- Después de 5 meses, los polluelos desnudos desarrollan un plumaje escaso y ya son capaces de soportar temperaturas bastante bajas.
En condiciones naturales, es probable que esta mutación sea letal y provoque una mortalidad del 100% de las aves. El ejemplo anterior muestra que la naturaleza de la manifestación de un gen semiletal puede depender en gran medida de las condiciones ambientales.
Los genes letales pueden ser dominantes o recesivos. Entre los primeros factores letales se descubrió el alelo que provocaba la coloración amarilla de los ratones. El gen del color amarillo es dominante (Y). Su alelo recesivo (y) en estado homocigoto provoca la aparición de una coloración negra. Al cruzar ratones amarillos entre sí se obtuvieron dos partes de ratones amarillos y una parte de ratones negros, es decir, el resultado fue una división de 2:1, y no de 3:1, como se desprende de la regla de Mendel. Resultó que todos los ratones adultos son heterocigotos (Yy). Al cruzarse entre sí deberían haber producido una parte de la descendencia homocigota para el color amarillo (IT), pero esta muere en el período embrionario, dos partes de los heterocigotos (Yy) serán amarillas y una parte de los homocigotos para el rasgo recesivo (yy) será negro. El esquema de cruce se ve así:
Del mismo modo, el color del pelaje gris se hereda en las ovejas Karakul (Sokolskie, Malich, etc.), el color platino en los zorros, la distribución de escamas en las carpas lineales, etc.
Los genes letales son en la mayoría de los casos recesivos y, por tanto, pueden permanecer latentes durante mucho tiempo.
Un animal completamente sano y de fenotipo normal puede ser portador de un gen letal, cuyo efecto se detecta sólo tras la transición a un estado homocigoto. Los genes letales suelen pasar a un estado homocigoto durante la endogamia. En la práctica de la cría de animales en la cría de caballos, hubo un caso de muerte de 25 potros entre el segundo y el cuarto día después del nacimiento por deformidad rectal - ausencia de ano (Atresia ani). Resultó que todos los sementales y yeguas que dieron a luz a potros tan anormales descendían del mismo semental. Era heterocigoto para el gen letal (LI). Inicialmente, este semental, cuando se cruzó con yeguas normales (LL), dio a luz a crías que eran normales en fenotipo, pero en términos de genotipo, la mitad de las crías eran normales (LL) y la otra mitad eran heterocigotas (LI), portando un inclinación recesiva (0 gen letal. En la consanguinidad de animales heterocigotos (Y x Y), aparecieron algunos potros, homocigotos para el gen letal (II), con deformidad rectal. Todos murieron.
Se conocen decenas de anomalías en los animales de granja, cuya aparición está asociada con mutaciones genéticas recesivas o dominantes. Estas anomalías se presentan en poblaciones individuales con diferente frecuencia, lo que depende de la velocidad del proceso de mutación, del sistema de cría del animal, etc. Conocimiento de las formas específicas de anomalías congénitas en animales de cada especie, así como de la frecuencia de su manifestación en razas individuales, es necesario para los especialistas veterinarios para la prevención selectiva de la propagación de patología genética.
Anomalías en el ganado. Las características biológicas de este tipo de animales son el pelo corto y una maduración relativamente tardía. Una vaca generalmente da a luz a una cría, que alcanza la madurez sexual y fisiológica solo al año y medio, por lo que el período entre el primer parto de una madre y una hija es en promedio de 5 años. Como resultado, la aparición de crías anormales en un rebaño puede reducir significativamente el nivel de reproducción y la intensidad de la selección genética del ganado. En el ganado se ha estudiado una amplia gama de shamalia congénita, determinada por genes letales, semiletales y subvitales. 46 anomalías están incluidas en la Lista Internacional de Defectos Letales bajo el código A (Tabla 43). La frecuencia relativa de tipos particulares de anomalías en cada raza o población puede variar. En la raza Kostroma, según nuestros datos, la anomalía genética de la cabeza registrada con mayor frecuencia es el acortamiento de la mandíbula (Tabla 44), en la raza Yaroslavl - sindactilia, en la raza Kholmogory - contracturas musculares, en la raza Negro y- Blanco - hernia umbilical. Las anomalías del sistema nervioso central fueron las más comunes (21%) en el ganado bovino en Alemania.
El segundo lugar en frecuencia de registro (14%) lo ocupó una anomalía compleja: una combinación de hernia umbilical con abdomen hendido y el feto en su conjunto. La frecuencia de anomalías, o el porcentaje de descendencia anormal con respecto al número total, dentro de poblaciones específicas también puede ser muy diferente y, según estimaciones promedio, no supera el 1%. Sin embargo, este indicador depende de la integridad y precisión del registro de anomalías. Así, en Alemania, después de organizar una contabilidad clara, llegaron a la conclusión de que la frecuencia de las anomalías se ha multiplicado varias veces. La pregunta es: ¿todas las anomalías son susceptibles de observación visual? Obviamente no todos. Así, en la raza Kostromá, la frecuencia media de todas las formas de anomalías durante un período de 12 años fue del 1,15%. La frecuencia de mortalidad general de la camada (terneros abortados, nacidos muertos, anormales, muertos sin defectos visibles) en esta granja fue del 10,2%. Una cierta proporción de esta mortalidad también está asociada con mutaciones genéticas que no causan defectos morfológicos, sino trastornos metabólicos y otras anomalías, cuya identificación sólo es posible mediante métodos especiales.
Los productores pueden desempeñar un papel especial en la propagación de anomalías genéticas tanto en el ganado como en otras especies animales. De cada padre, con inseminación artificial, se pueden obtener cientos y miles de crías al año. Así, de un toro se obtuvieron 100 mil terneros en el extranjero. Si dicho padre resulta ser portador de una mutación genética, ésta se propagará rápidamente por toda la raza. A continuación se muestran algunos ejemplos de los numerosos hechos descritos en la literatura. Como resultado del uso intensivo del toro Príncipe Adolf, traído a Suecia, y la posterior endogamia espontánea con él, la frecuencia de falta de pelo en algunas manadas suecas superó el 5%. La misma situación se produjo en Suecia tras la importación del toro Gallus, que resultó ser portador heterocigoto del gen que provoca la ausencia de extremidades.
En la descendencia de toros individuales Blanco y Negro y Charolais en EE. UU. y Alemania, se registraron casos de nacimiento de terneros enanos con una frecuencia de 23,3 y 22,2%, respectivamente. En la antigua En Checoslovaquia, al examinar a los descendientes de 166 toros, se encontró que 43 de ellos eran portadores de genes letales. En un toro, portador de la anomalía dominante "labio hendido", el defecto se manifestó en el 44% de los toros y en el 71% de las novillas de su descendencia.
En la raza Kostromá analizamos la propagación del acortamiento de la mandíbula inferior y la apariencia de pug en el toro Burkhan, que también tenía crías defectuosas en su descendencia; sus hijos, nietos, bisnietos y descendientes femeninas también dieron descendencia anormal (Fig. 57). La mayoría de los terneros se obtienen por endogamia y apareamiento de padres con un fenotipo normal y un ancestro común. Por tanto, podemos concluir que esta anomalía tiene un modo de herencia recesivo. La figura muestra que el mayor número de terneros defectuosos se registró en la descendencia del toro Zheton 3501 (nieto del toro Burkhan) cuando se utilizó en una granja comercial, donde algunas de las vacas tenían el mismo gen recesivo en su genotipo.
Anomalías en cerdos. La Lista Internacional de Defectos Letales en Porcinos incluye 18 anomalías genéticas. La mayor parte de ellos son causados por genes autosómicos recesivos (Tabla 45). Las anomalías genéticas pueden desempeñar un papel importante en la patología de los cerdos. Veamos algunos de estos ejemplos. En España, un estudio de 23.449 lechones de 2.399 camadas obtenidas de verracos de las razas Duroc, Yorkshire, Hampshire y White Chester reveló, respectivamente, 6,21; 6,02; 9,66; 2^62% de camadas anormales.
Según Olivier (1979), 7 anomalías genéticas de la piel, 17 del esqueleto, 3 de los ojos, 13 del sistema neuromuscular, 6 de la sangre, 6 del metabolismo hormonal, 5 del sistema digestivo y 9 del sistema genitourinario se han descrito en cerdos. Las principales anomalías fueron criptorquidia, hernias, pseudohermafroditismo, etc. El autor del estudio cree que estas anomalías son el resultado de la acción de un gen en diferentes etapas de la formación del embrión.
En Dinamarca, se estudiaron 6.669 lechones muertos de 2.936 camadas durante un período de dos años para determinar la naturaleza y frecuencia de las anomalías congénitas. Se detectaron diversas anomalías en el 1,4% de los lechones nacidos o en el 6,2% de los sacrificados antes del destete. Durante un examen post mortem, se encontró que el 25,9% de los lechones anormales tenían válvulas subdesarrolladas, falta de cierre del ano, estenosis subaórtica, ectopia del corazón y otros defectos del sistema cardiovascular. Se encontraron diversos trastornos del desarrollo del sistema motor en el 23,4% de los lechones. Se detectaron anomalías del sistema nervioso central en el 5,9% de los lechones, incluido cerebro bifurcado e hidrocele cerebral. En el 30% de los lechones se encontró un crecimiento excesivo del recto, el intestino delgado o su desarrollo incompleto, y diversas hernias y ascitis, en el 6,8%. “labio hendido”, paladar hendido, rinocefalitis y otras anomalías de la parte facial de la cabeza se encontraron en el 6,1%; hermafroditismo, fisura ureteral, hidrocele de riñón y uretra, en el 1,7% de los lechones. Estas anomalías aparecieron en la descendencia de toros individuales durante la endogamia, lo que indica la naturaleza hereditaria de su aparición.
Fridin y Newman obtuvieron pruebas muy convincentes de la naturaleza hereditaria de la criptorquidia en cerdos. Según sus datos, en Canadá, la criptorquidia unilateral y bilateral se observa anualmente en el 1-2% de todos los verracos que ingresan al mercado. Los autores cruzaron criptorquídeas con sus madres y hermanas completas. Los descendientes de tales cruces se aparearon entre sí. Como resultado de dicha selección y selección, la frecuencia de criptorquidia en animales de experimentación de la raza Yorkshire aumentó a un promedio del 42,9%, y especialmente cuando se utilizaron dos productores. Al examinar lechones en los Estados Unidos durante un año, se encontraron alrededor de 400 mil animales con una hernia escrotal.
Las observaciones muestran que la causa de la alteración de la fertilidad en los verracos es a menudo la hipoplasia de los testículos. La frecuencia de esta anomalía, según investigadores alemanes, fue del 19,6%. Se dejaron 30 de estos jabalíes para su reproducción.
Va, cada una de ellas tenía de 4 a 40 reinas (un total de 439 cabezas), pero solo cuatro de ellas dieron a luz a descendencia. El análisis mostró que las formas patológicas de esperma en estos verracos son del 80 al 100%. Los 30 animales anormales tenían ancestros comunes, lo que indica la naturaleza hereditaria de la hipoplasia testicular y los defectos de la espermiogénesis.
La presencia de pezones con cráteres en los cerdos es uno de los defectos graves, ya que los lechones no reciben leche de ellos. Según el Instituto Bávaro de Ganadería (Alemania), la frecuencia de esta anomalía en las razas locales alemanas fue del 6,6%. Como señalaron P. N. Kudryavtsev et al. (MVA), el número de cerdos con pezones con cráter inactivos ha aumentado en los últimos años. El número de estas tetinas varía de 1 a 8. Los lechones que reciben tetinas con cráter mueren.
La crateridad es un rasgo causado por un gen autosómico recesivo. Esto fue probado experimentalmente por P. N. Kudryavtsev et al. Habiendo identificado previamente verracos y primerizas portadores de anomalías (KchKch), individuos normales pero heterocigotos (KchKch), que producen lechones con cráter en su descendencia, y cerdos homocigotos normales (KhKch), los autores realizaron cruces entre estos grupos de animales. . En la primera variante, se cruzaron 27 madres homocigotas normales con 15 verracos. Los 258 descendientes eran normales. En la segunda opción, donde uno de los padres era homocigoto (KchKch) y el otro heterocigoto (Kchkch), todos los lechones también eran normales. En la tercera variante, se cruzaron 13 verracos heterocigotos con 16 madres heterocigotas. De los 168 lechones nacidos, 39 (23,2%) tenían pezones con cráter. Y finalmente, en la cuarta opción, uno de los padres era homocigoto y el otro heterocigoto. Produjeron 170 lechones, de los cuales 86 (50,5%) tenían pezones normales y 84 (49,5%) tenían pezones con cráter. Los resultados de este experimento demuestran el modo recesivo de herencia del cráter del pezón en los cerdos.
Anomalías en ovejas. Se han descrito alrededor de 90 anomalías congénitas en ovejas. Según Dennis y Leipold, la mayoría de los defectos genéticos conocidos en ovejas son causados por un modo de herencia autosómico recesivo monogénico (Tabla 46). Los defectos craneofaciales más comunes en esta especie de animales son defectos craneofaciales, especialmente agnatia, así como curvatura de las extremidades anteriores, microagnatia, hermafroditismo, criptorquidia, hipospadiasis, prognatia, atresia anal, microtia, entropía, tortícolis, politelia, artrogriposis. El análisis mostró que el 55,4% de los defectos estaban relacionados con el sistema musculoesquelético, 12,7 - con el sistema digestivo, 9,7 - con el sistema cardiovascular, 7,1 - con el sistema urogenital, 6 - con el sistema nervioso central, 3,5 - con anomalía de los ligamentos, 3,2 - al abdominal, 1,5% - al sistema endocrino. Aunque la frecuencia de defectos individuales es baja, la contribución acumulativa de todas las anomalías puede causar daños a las explotaciones. En Nueva Zelanda, un país con una cría de ovejas desarrollada, la incidencia de defectos letales fue de aproximadamente el 1% de los corderos muertos. En EE.UU., se observaron defectos letales en las dos primeras semanas de vida en el 11,4% de los corderos.
La tasa media de mortalidad embrionaria en ovejas es del 20%. Esto indica que muchos genes letales no identificables pueden estar activos durante este período.
Las anomalías genéticas individuales en las ovejas pueden estar muy extendidas. Así, en Bulgaria, en los rebaños de ovejas merinas, se observó una alta mortalidad de los corderos en el período posnatal temprano. Ocurrió a raíz de que los corderos no recibían leche por defectos en la ubre de sus madres: desde hipoplasia con pequeños restos de tejido glandular hasta su total ausencia. La frecuencia de esta anomalía hereditaria en diferentes rebaños osciló entre el 6 y el 40%.
En las ovejas, a menudo se observaba criptorquidia, que se combinaba con una característica sexual secundaria como la falta de cuernos. Los carneros sin cuernos tienen baja fertilidad. A través de una estricta selección genética, fue posible crear un tipo de carneros sin cuernos con fertilidad normal, sin embargo, entre ellos también se encuentran individuos anormales.
Anomalías en las aves. Las aves, principalmente las gallinas, han sido las más estudiadas en relación con la genética de las anomalías. La Lista Internacional de Defectos Letales incluye 45 anomalías en pollos, 6 en pavos y 3 en patos. Las anomalías del pico más comunes son (pico de loro, pico cruzado). Su frecuencia, según William et al., es del 1,1% de las pérdidas durante la incubación de huevos de gallinas White Leghorn y Rhode Island. Las anomalías en el pico también son comunes en los patos.
Max Gibbon y Shakelfurd describieron una anomalía al cruzar leghorns blancos con buttercamps y bantams y su posterior reproducción "en sí mismos": la polidactilia. Además, se observaron sindactilia y patas emplumadas en los pollos. La frecuencia del síndrome fue del 16,8%. Al cruzar un gallo F2 anormal con gallinas fenotípicamente normales, se observó una división: mitad individuos normales y mitad individuos anormales (1:1). Se ha establecido que este complejo de rasgos está controlado por un gen autosómico, que tiene un efecto semiletal, ya que la supervivencia embrionaria y postembrionaria de los pollos anormales es muy baja.
Anomalías en los caballos. De las anomalías hereditarias en los caballos, 10 están incluidas en la Lista Internacional de Defectos Letales. Entre ellas se encuentran 3 anomalías del esqueleto, 2 del sistema reproductivo, 2 de los riñones y músculos, una anomalía de cada uno de los intestinos, del sistema nervioso y de los órganos de la visión.
En los caballos de razas de tiro, la atresia de colon es más común. La prevalencia de esta anomalía se observó en la descendencia del semental Superba de la raza Percheron. También se ha descrito en potros de una raza de pura raza de origen africano oriental. Se ha registrado epiteliogénesis imperfecta en razas de tiro. La ataxia, llamada Oldenburg, se encontró en potros de Oldenburg en Alemania. Se extendió en la línea 9. Una de las anomalías registradas con más frecuencia en los caballos es la hernia umbilical. Se encuentra en razas ligeras y pesadas.
Se conocen varias otras anomalías genéticas y hereditarias-ambientales en los caballos. Así, en Estados Unidos, los caballos de varias razas tienen potros con una peculiar mancha blanca, llamada “overo”. Al cruzar caballos del tipo “overo”, nacen potros con piel rosada, en los que se observa hipoplasia del tracto intestinal e isoeritrolisis, además de cólicos que provocan la muerte.
En Inglaterra, se registraron y estudiaron animales con problemas de coordinación de movimientos (la "enfermedad del bamboleo") en caballos mestizos de raza de montar. Se ha establecido una predisposición genética a esta anomalía.
Se ha demostrado la heredabilidad de las dermatosis de las extremidades en caballos. Los defectos con predisposición hereditaria son la inflamación crónica deformante de la articulación del corvejón, la llamada "pata de muleta" en los potros, así como la inflamación crónica aséptica del bloque coronal de los cascos, que se observa principalmente en los caballos de carreras y carreras. caballos.









- Galletas de mantequilla de limón Cómo hacer galletas de mantequilla de limón
- Receta de ensalada Yeralash con ternera
- Salmón rosado al horno con patatas
- Cómo cocinar maleza en casa: recetas deliciosas y fáciles
- Basturma casera - las mejores recetas
- Cómo organizar un escritorio según el Feng Shui por dinero
- Las conspiraciones contra un rival devolverán la paz a la familia
- Apuntes sobre la alfabetización en el grupo preparatorio “Viajes espaciales”
- Oficial Sergei Rybakov: “El tiempo es lo que le dedicamos
- Dolor en el lado izquierdo desde la espalda.
- En coche - del estado
- Finanzas en economía. Sistema bancario. Finanzas en economía Presentación estudios sociales 11º grado finanzas en economía
- Presentación sobre el tema de las finanzas en la economía.
- Origen e historia del pueblo ávar.
- Dispositivos médicos para el tratamiento de las articulaciones en el hogar Dispositivo de fisioterapia ultrasónico doméstico para el tratamiento de las articulaciones
- Precios unitarios territoriales
- Levantamiento de Kronstadt ("rebelión") (1921) Represión del levantamiento de Kronstadt
- Sistema taoísta. L. BingSecretos del amor. Práctica taoísta para mujeres y hombres. Sistema "Tao Universal"
- Qigong: práctica china para fortalecer el cuerpo
- Sociedad Oed para la evangelización infantil